

Regulation of phosphorylation of the GluR1 AMPA receptor in the neostriatum by dopamine and psychostimulants in vivo
- PMID: 10844017
- PMCID: PMC6772453
- DOI: 10.1523/JNEUROSCI.20-12-04480.2000
Regulation of phosphorylation of the GluR1 AMPA receptor in the neostriatum by dopamine and psychostimulants in vivo
Abstract
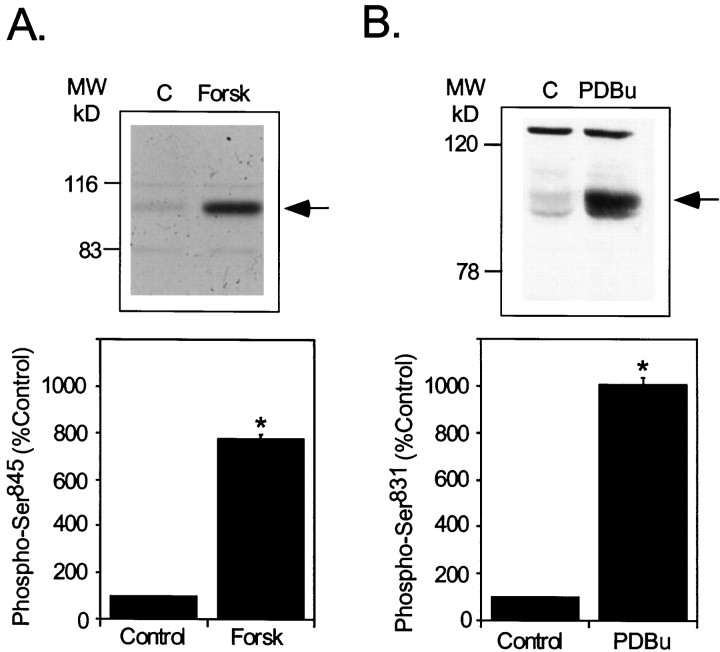
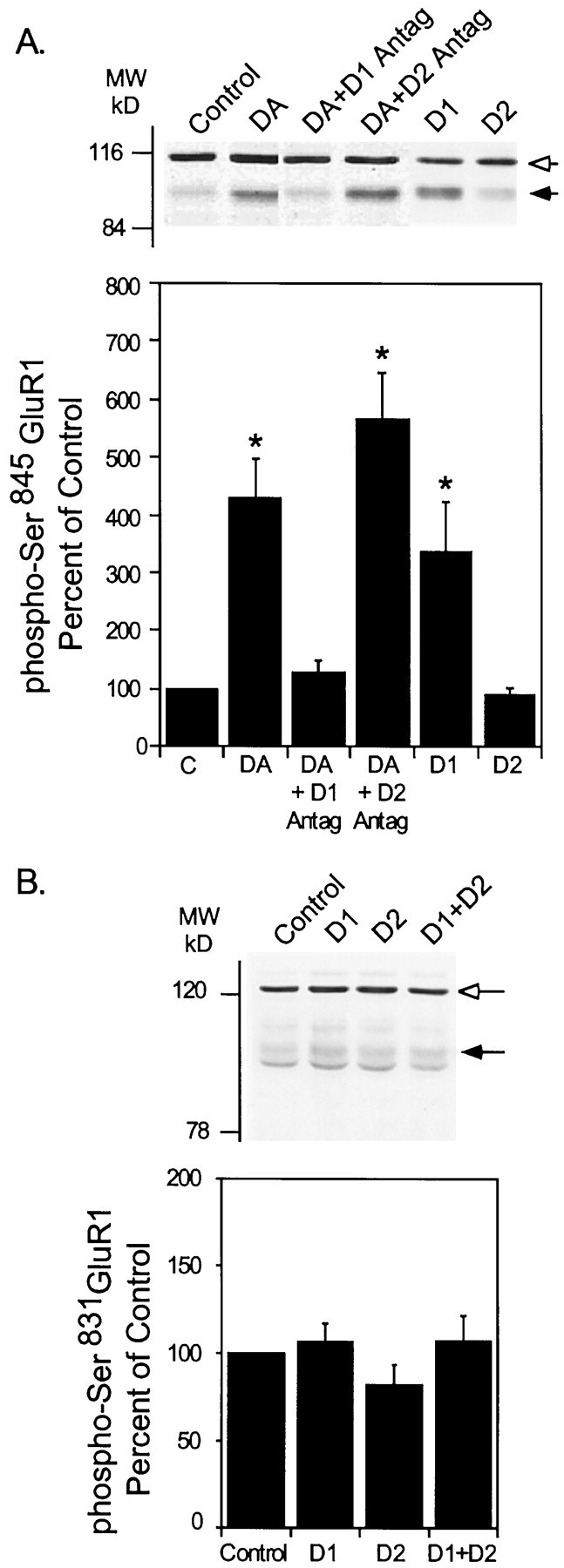
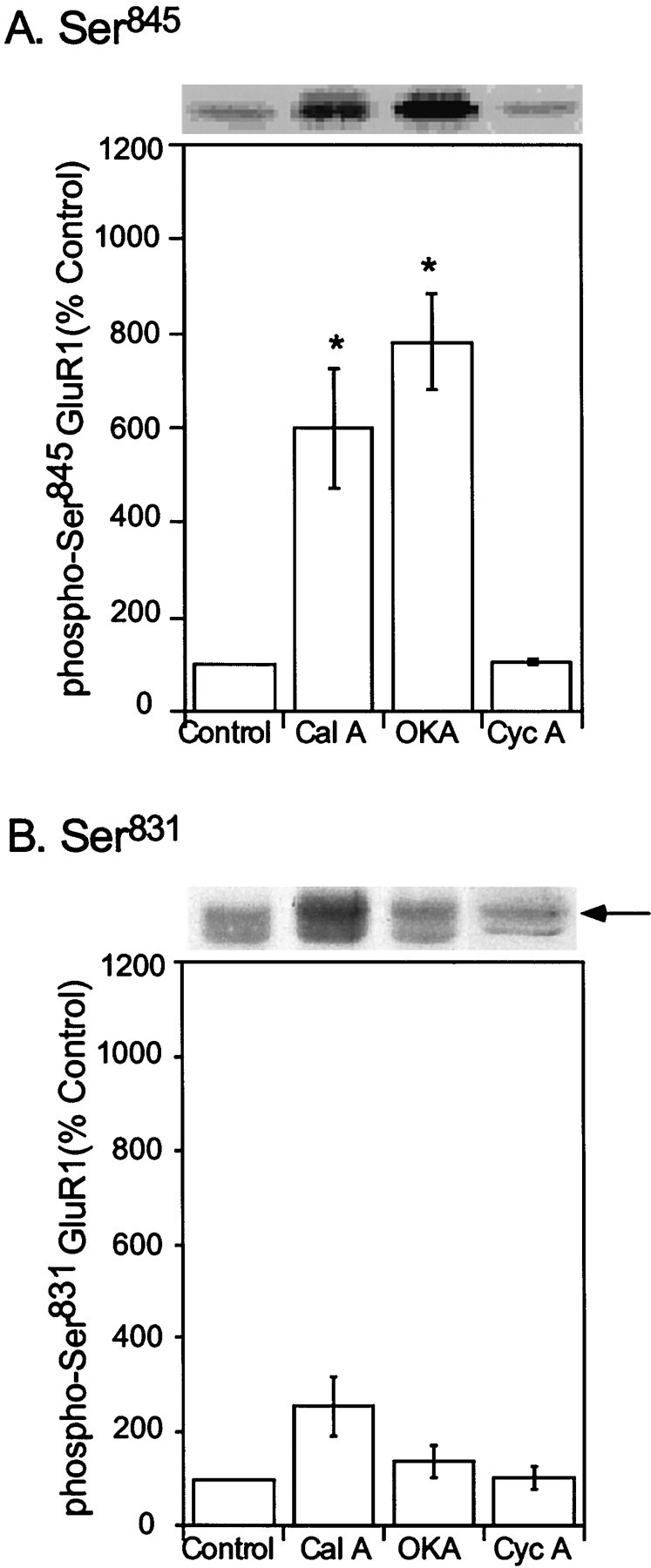
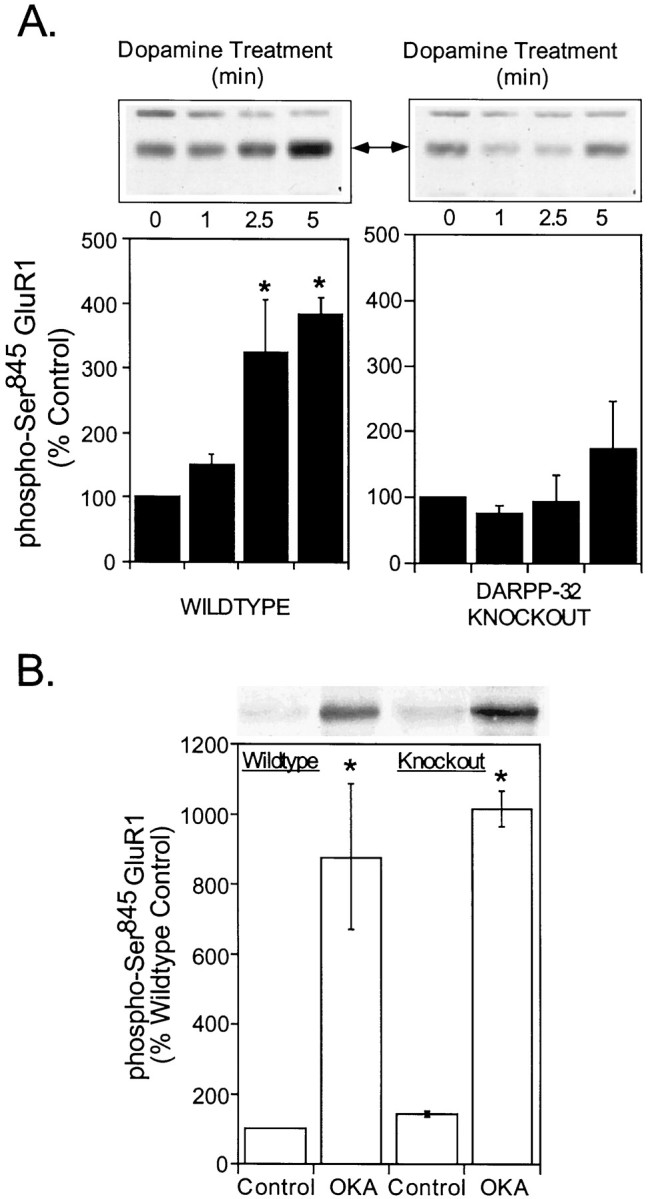
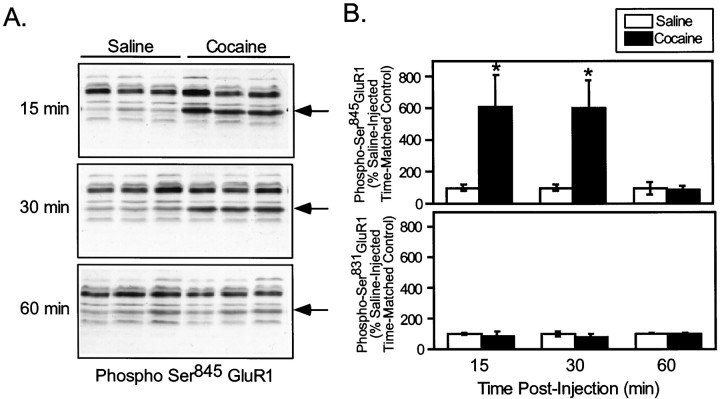
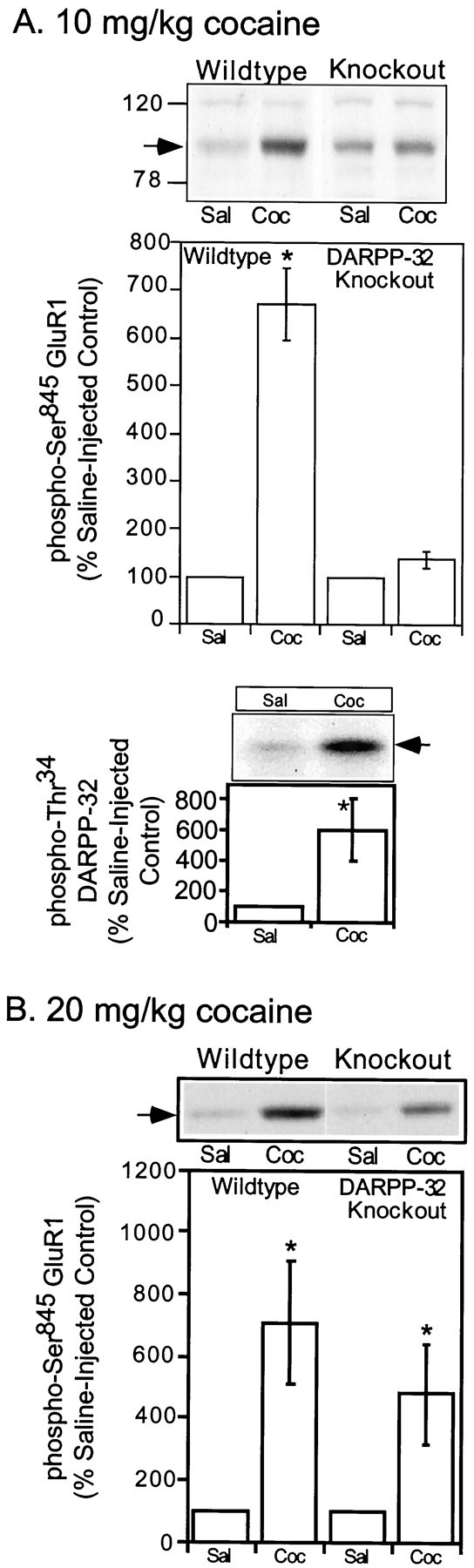
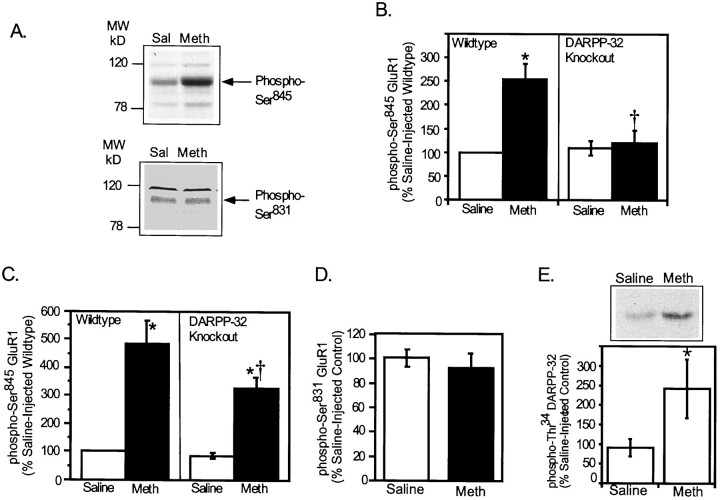
The activation of cAMP-dependent protein kinase regulates the physiological activity of AMPA-type glutamate receptors. In this study, phosphorylation of the AMPA receptor subunit GluR1 at Ser(845) was increased in neostriatal slices by activation of D1-type dopamine receptors and by inhibitors of protein phosphatase 1/protein phosphatase 2A. In contrast, Ser(831), a residue which, when phosphorylated by protein kinase C or calcium/calmodulin-dependent kinase II, increases AMPA receptor channel conductance, was unaffected by either D1 or D2 receptor agonists in neostriatal slices. The phosphorylation of Ser(845), but not Ser(831), was strongly increased in neostriatum in vivo in response to the psychostimulants cocaine and methamphetamine. The effects of dopamine and psychostimulants on the phosphorylation of GluR1 were attenuated in dopamine and cAMP-regulated phosphoprotein M(r) 32 kDa (DARPP-32) knock-out mice. These results identify DARPP-32 and AMPA-type glutamate receptors as likely essential cellular effectors for psychostimulant actions.
Figures
References
-
- Barria A, Muller D, Derkach V, Griffith LC, Soderling TR. Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science. 1997;276:2042–2045. - PubMed
-
- Bibb JA, Snyder GL, Nishi A, Yan Z, Meijer L, Fienberg AA, Tsai L-H, Kwon YT, Girault J-A, Czernik AJ, Huganir RL, Hemmings HC, Jr, Nairn AC, Greengard P. Phosphorylation of DARPP-32 by Cdk5 modulates dopamine signalling in neurons. Nature. 1999;402:669–671. - PubMed
-
- Bliss TVP, Collingridge GL. Synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases