

Local disparity not perceived depth is signaled by binocular neurons in cortical area V1 of the Macaque
- PMID: 10844045
- PMCID: PMC6772469
- DOI: 10.1523/JNEUROSCI.20-12-04758.2000
Local disparity not perceived depth is signaled by binocular neurons in cortical area V1 of the Macaque
Abstract
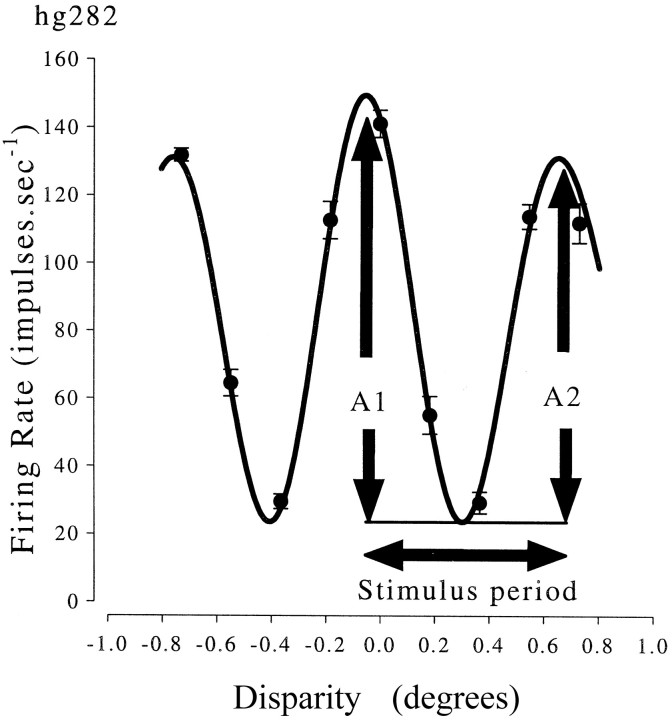
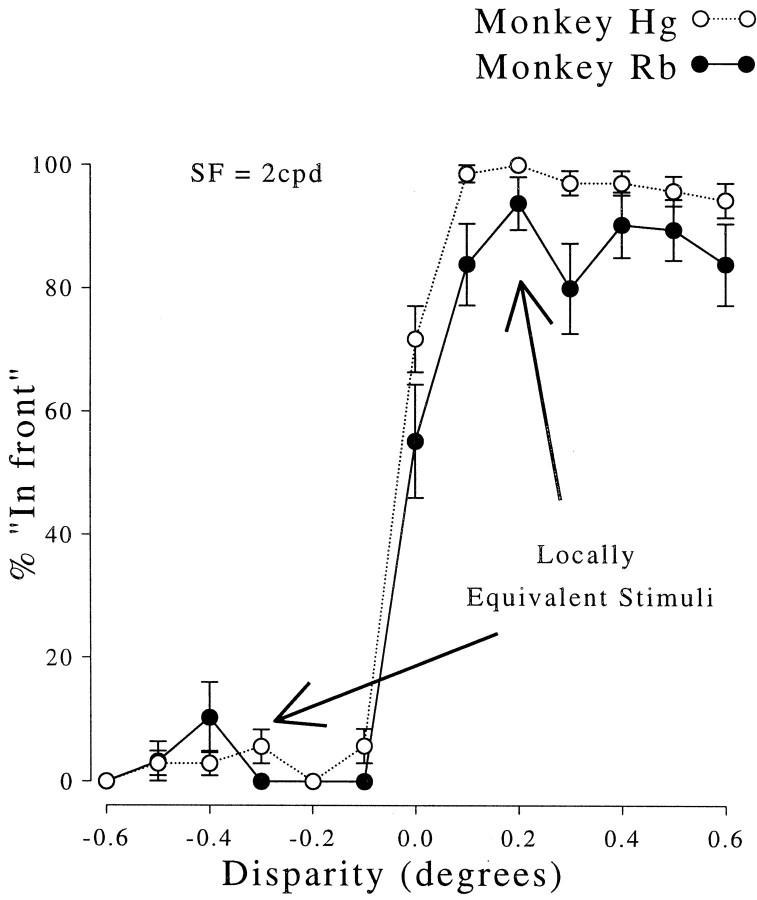
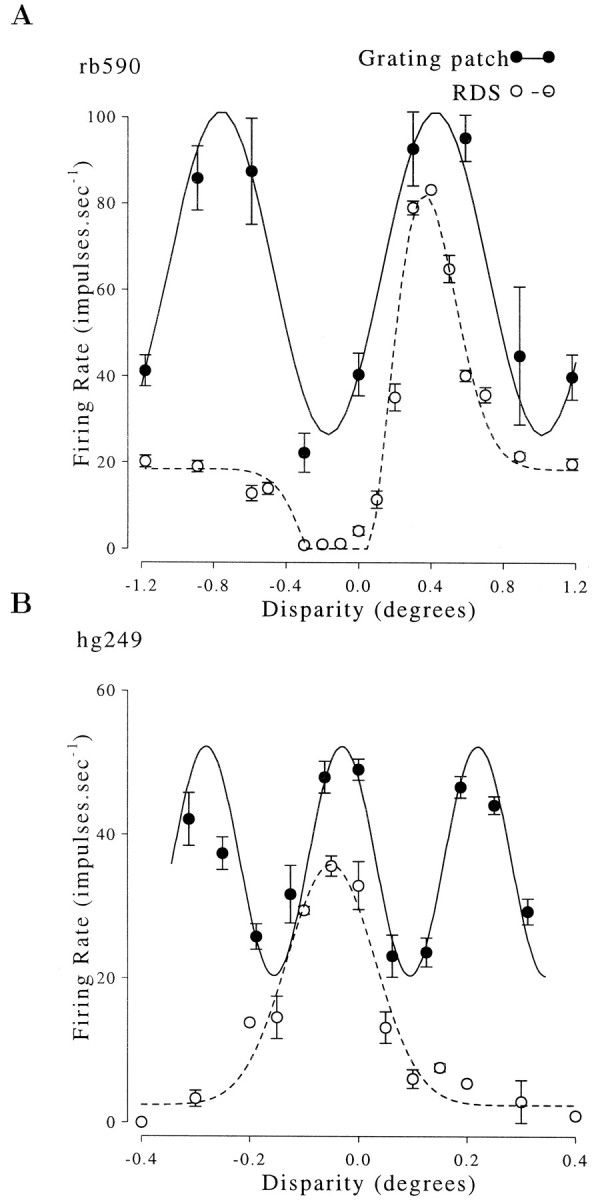
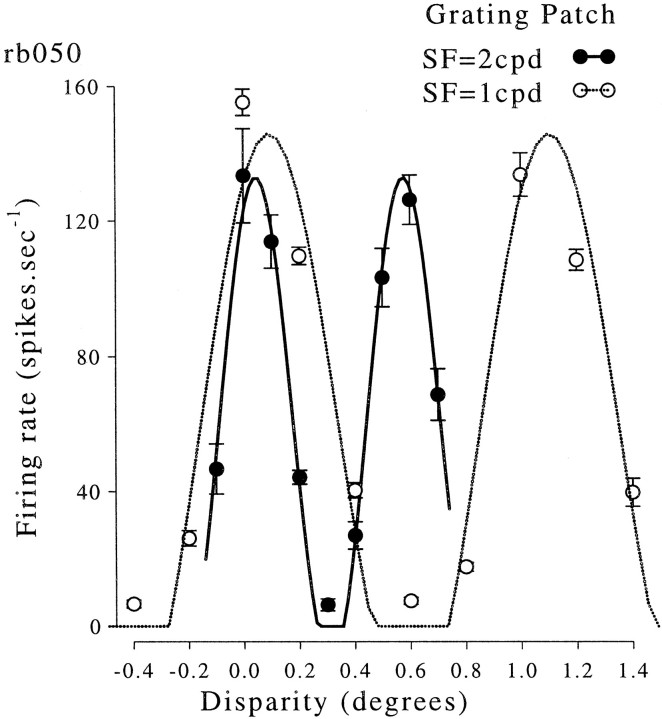
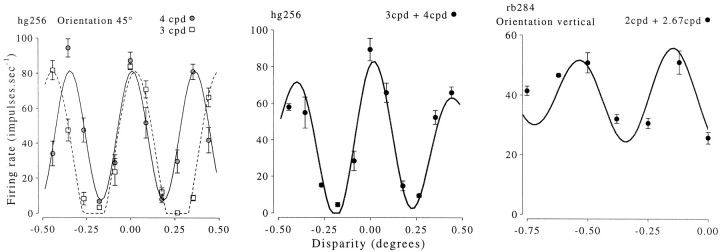
Binocular neurons that are closely related to depth perception should respond selectively for stimuli eliciting an appropriate depth sensation. To separate perceived depth from local disparity within the receptive field, sinusoidal luminance gratings were presented within a circular aperture. The disparity of the aperture was coupled to that of the grating, thereby rendering unambiguous the psychophysical matching between repeating cycles of the grating. In cases in which the stimulus disparity differs by one horizontal period of the grating, the portion of the grating that locally covers a receptive field is binocularly identical, but the depth sensation is very different because of the aperture. For 117 disparity-selective V1 neurons tested in two monkeys, the overwhelming majority responded equally well to configurations that were locally identical but led to different perceptions of depth. Because the psychophysical sensation is not reflected in the firing rate of V1 neurons, the signals that make stereo matches explicit are most likely elaborated in extrastriate cortex.
Figures

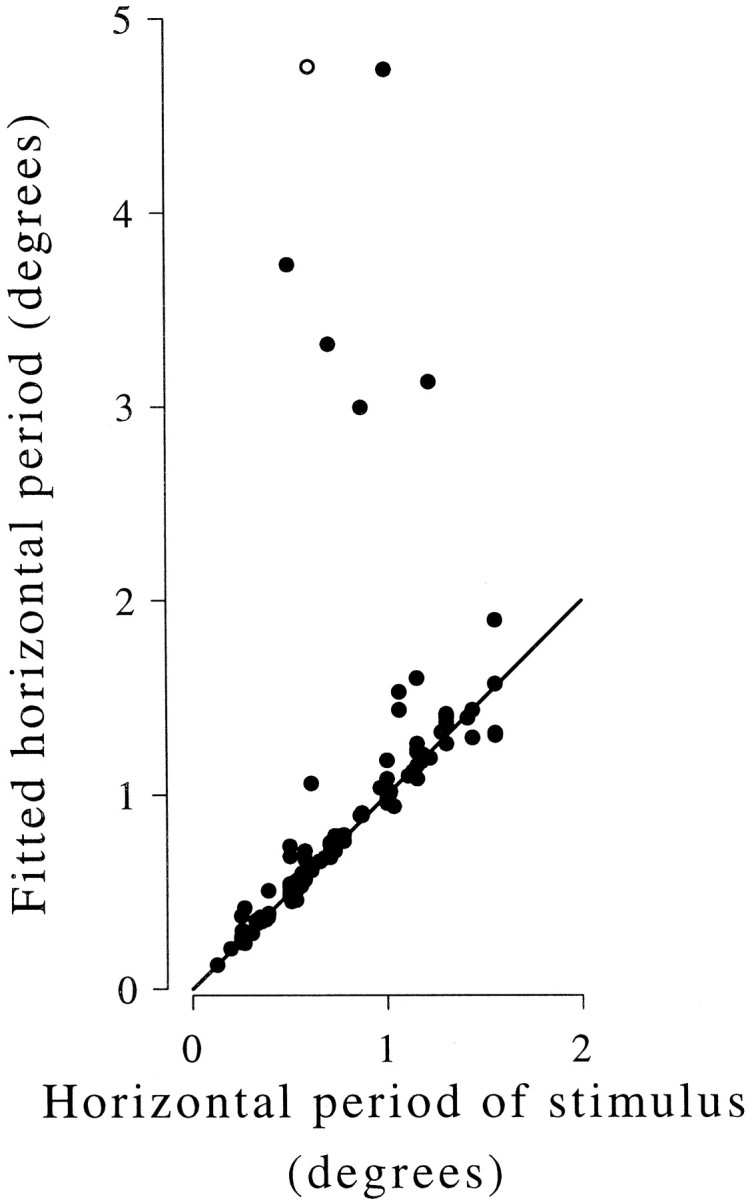
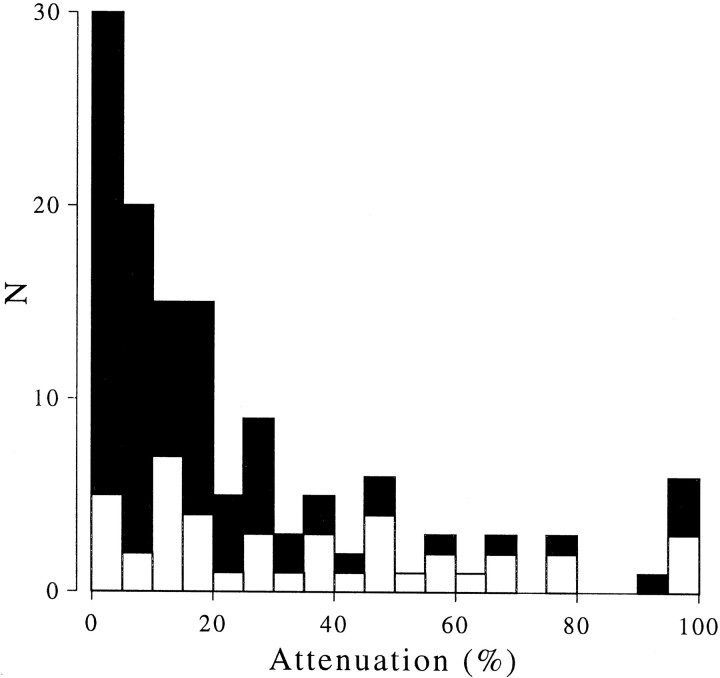
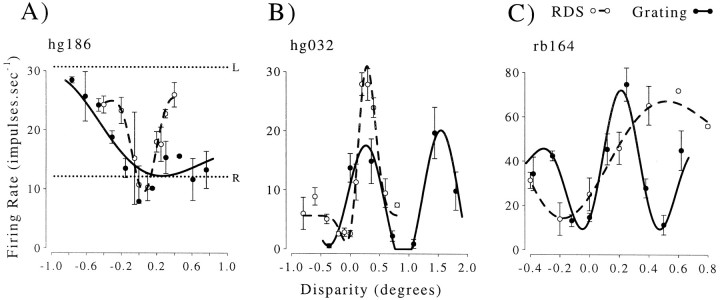

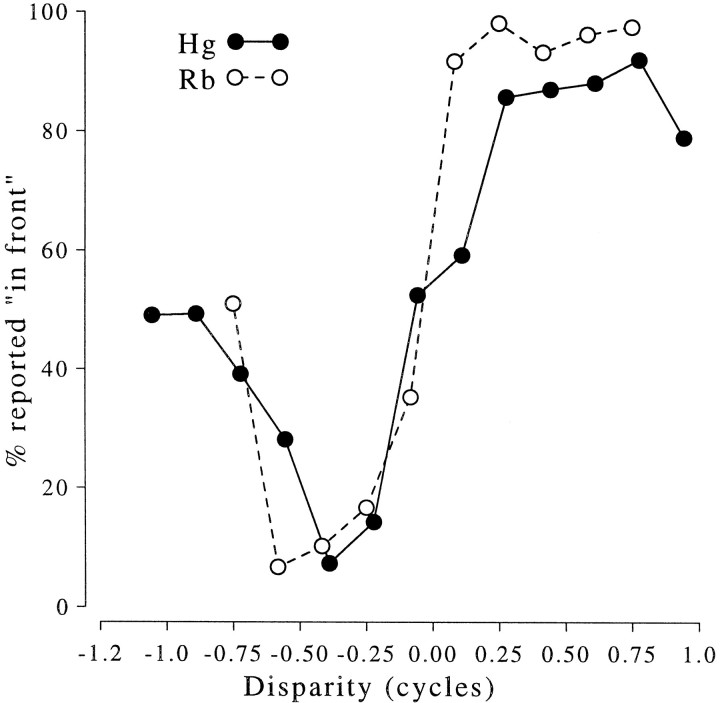
References
-
- Armitage P, Berry G. Statistical methods in medical research. Blackwell; Oxford: 1994.
-
- Britten KH, Shadlen MN, Newsome WT, Movshon JA. Responses of neurons in macaque MT to stochastic motion signals. Vis Neurosci. 1993;10:1157–1169. - PubMed
-
- Cogan A, Lomakin A, Rossi A. Depth in anticorrelated stereograms. Vision Res. 1993;33:1959–1975. - PubMed
-
- Cumming BG, Parker AJ. Responses of primary visual cortical neurons to binocular disparity without the perception of depth. Nature. 1997;389:280–283. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources