

Reovirus protein sigmaNS binds in multiple copies to single-stranded RNA and shares properties with single-stranded DNA binding proteins
- PMID: 10846075
- PMCID: PMC112090
- DOI: 10.1128/jvi.74.13.5939-5948.2000
Reovirus protein sigmaNS binds in multiple copies to single-stranded RNA and shares properties with single-stranded DNA binding proteins
Abstract
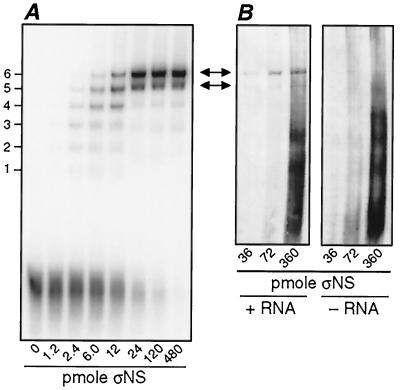
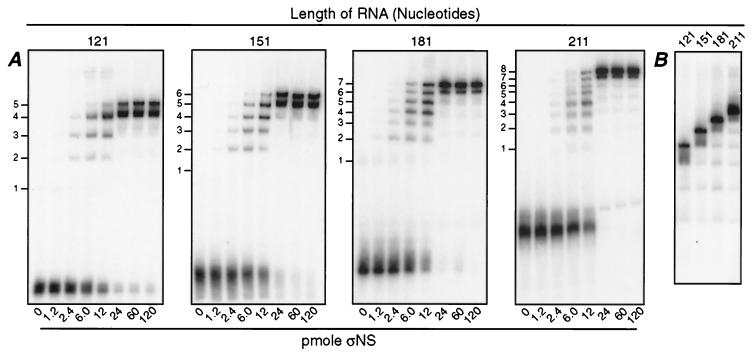
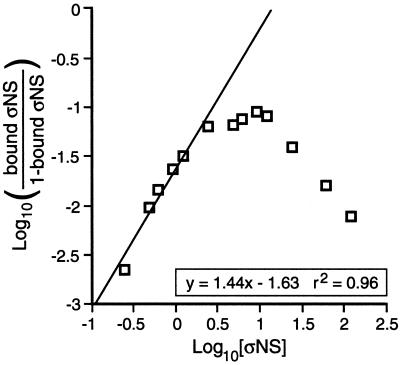
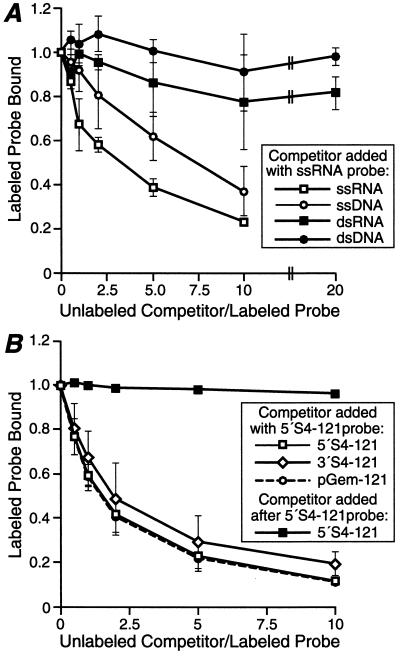
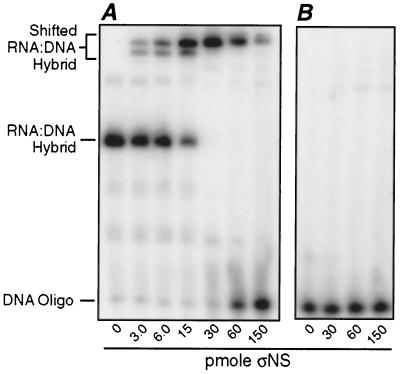
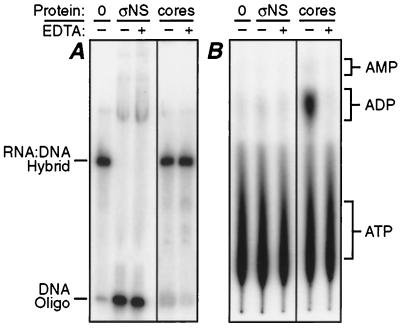
Reovirus nonstructural protein sigmaNS interacts with reovirus plus-strand RNAs in infected cells, but little is known about the nature of those interactions or their roles in viral replication. In this study, a recombinant form of sigmaNS was analyzed for in vitro binding to nucleic acids using gel mobility shift assays. Multiple units of sigmaNS bound to single-stranded RNA molecules with positive cooperativity and with each unit covering about 25 nucleotides at saturation. The sigmaNS protein did not bind preferentially to reovirus RNA over nonreovirus RNA in competition experiments but did bind preferentially to single-stranded over double-stranded nucleic acids and with a slight preference for RNA over DNA. In addition, sigmaNS bound to single-stranded RNA to which a 19-base DNA oligonucleotide was hybridized at either end or near the middle. When present in saturative amounts, sigmaNS displaced this oligonucleotide from the partial duplex. The strand displacement activity did not require ATP hydrolysis and was inhibited by MgCl(2), distinguishing it from a classical ATP-dependent helicase. These properties of sigmaNS are similar to those of single-stranded DNA binding proteins that are known to participate in genomic DNA replication, suggesting a related role for sigmaNS in replication of the reovirus RNA genome.
Figures
Similar articles
-
Reovirus Nonstructural Protein σNS Acts as an RNA Stability Factor Promoting Viral Genome Replication.J Virol. 2018 Jul 17;92(15):e00563-18. doi: 10.1128/JVI.00563-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29769334 Free PMC article.
-
Identification and characterization of RNA-binding activities of avian reovirus non-structural protein sigmaNS.J Gen Virol. 1998 Jun;79 ( Pt 6):1411-3. doi: 10.1099/0022-1317-79-6-1411. J Gen Virol. 1998. PMID: 9634083
-
Reovirus Nonstructural Protein σNS Recruits Viral RNA to Replication Organelles.mBio. 2021 Aug 31;12(4):e0140821. doi: 10.1128/mBio.01408-21. Epub 2021 Jul 6. mBio. 2021. PMID: 34225484 Free PMC article.
-
Reovirus sigma 3 protein: dsRNA binding and inhibition of RNA-activated protein kinase.Curr Top Microbiol Immunol. 1998;233(Pt 1):185-96. doi: 10.1007/978-3-642-72092-5_9. Curr Top Microbiol Immunol. 1998. PMID: 9599927 Review. No abstract available.
-
Function, Architecture, and Biogenesis of Reovirus Replication Neoorganelles.Viruses. 2019 Mar 21;11(3):288. doi: 10.3390/v11030288. Viruses. 2019. PMID: 30901959 Free PMC article. Review.
Cited by
-
Reovirus sigma NS and mu NS proteins form cytoplasmic inclusion structures in the absence of viral infection.J Virol. 2003 May;77(10):5948-63. doi: 10.1128/jvi.77.10.5948-5963.2003. J Virol. 2003. PMID: 12719587 Free PMC article.
-
Carboxyl-proximal regions of reovirus nonstructural protein muNS necessary and sufficient for forming factory-like inclusions.J Virol. 2005 May;79(10):6194-206. doi: 10.1128/JVI.79.10.6194-6206.2005. J Virol. 2005. PMID: 15858004 Free PMC article.
-
Virus-mediated compartmentalization of the host translational machinery.mBio. 2014 Sep 16;5(5):e01463-14. doi: 10.1128/mBio.01463-14. mBio. 2014. PMID: 25227463 Free PMC article.
-
Sequestration and protection of double-stranded RNA by the betanodavirus b2 protein.J Virol. 2006 Jul;80(14):6822-33. doi: 10.1128/JVI.00079-06. J Virol. 2006. PMID: 16809288 Free PMC article.
-
Identification and characterization of the helix-destabilizing activity of rotavirus nonstructural protein NSP2.J Virol. 2001 May;75(10):4519-27. doi: 10.1128/JVI.75.10.4519-4527.2001. J Virol. 2001. PMID: 11312322 Free PMC article.
References
-
- Antczak J B, Chmelo R, Pickup D J, Joklik W K. Sequence at both termini of the 10 genes of reovirus serotype 3 (strain Dearing) Virology. 1982;121:307–319. - PubMed
-
- Antczak J B, Joklik W K. Reovirus genome segment assortment into progeny genomes studied by the use of monoclonal antibodies directed against reovirus proteins. Virology. 1992;187:760–776. - PubMed
-
- Beckman M T L, Kirkegaard K. Site size of cooperative single-stranded RNA binding by poliovirus RNA-dependent RNA polymerase. J Biol Chem. 1998;273:6724–6739. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
