

Involvement of p21(Waf1/Cip1) in protein kinase C alpha-induced cell cycle progression
- PMID: 10848585
- PMCID: PMC85854
- DOI: 10.1128/MCB.20.13.4580-4590.2000
Involvement of p21(Waf1/Cip1) in protein kinase C alpha-induced cell cycle progression
Abstract
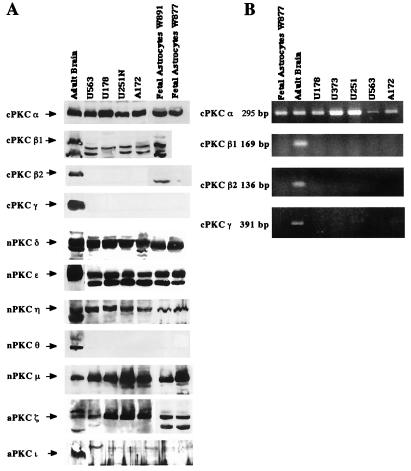
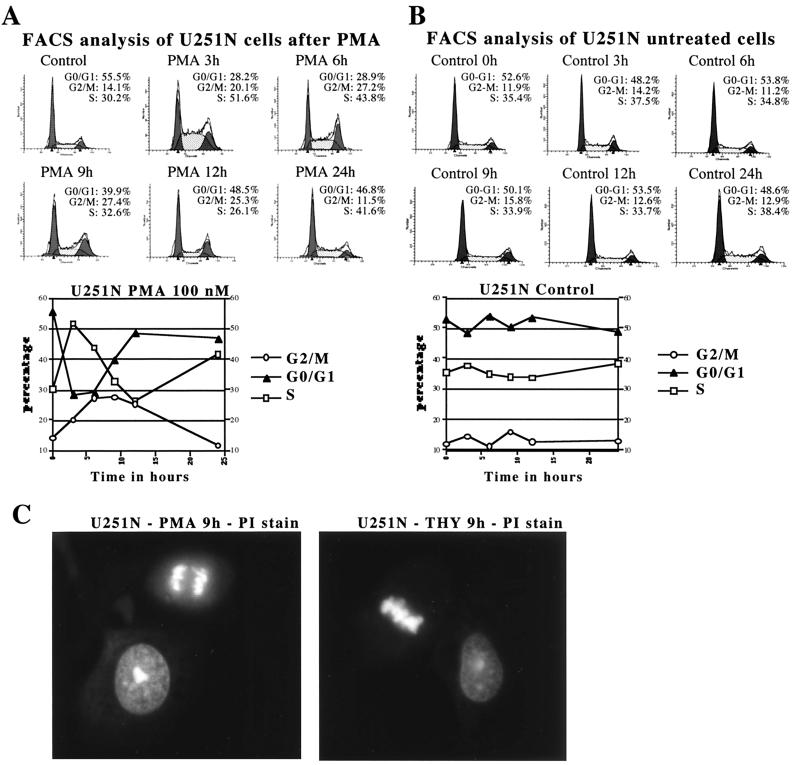
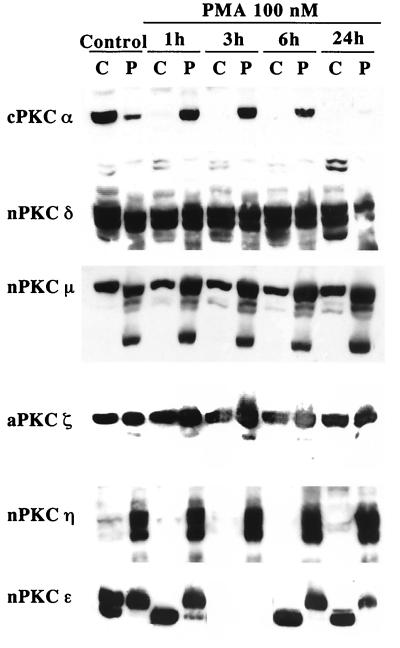
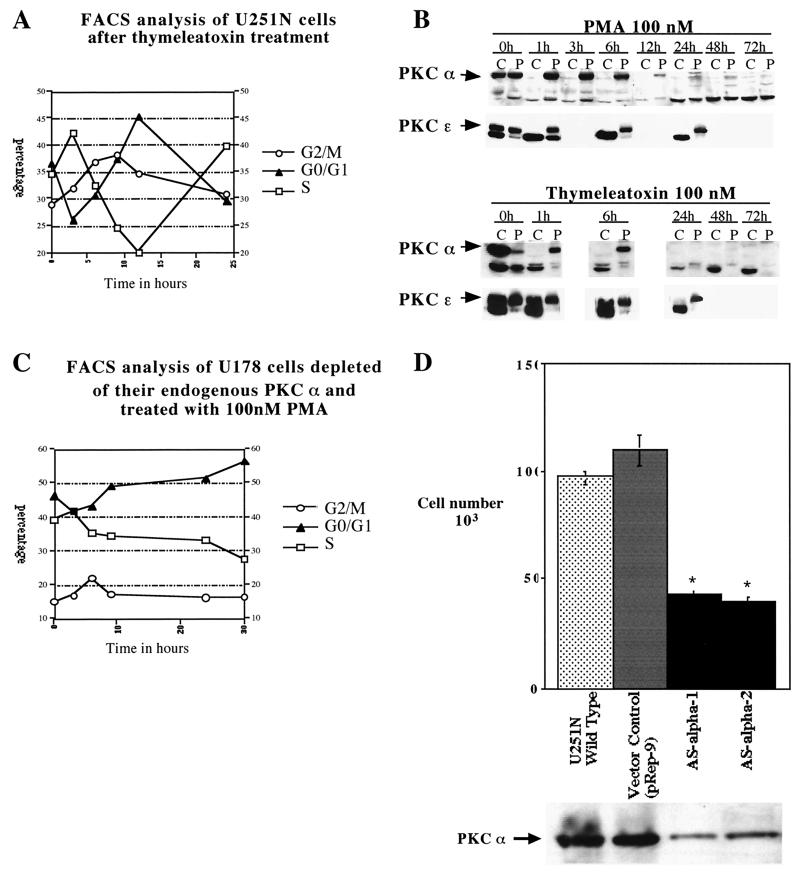
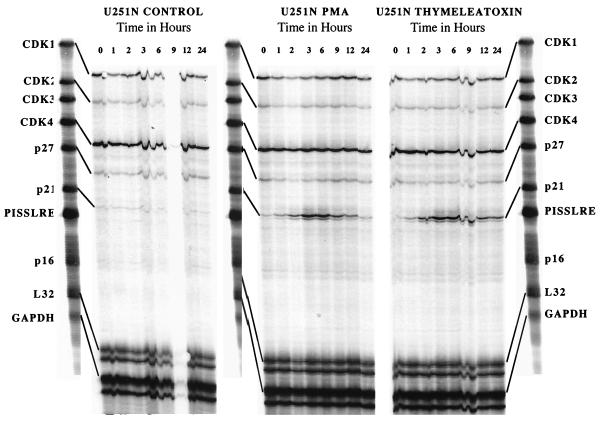
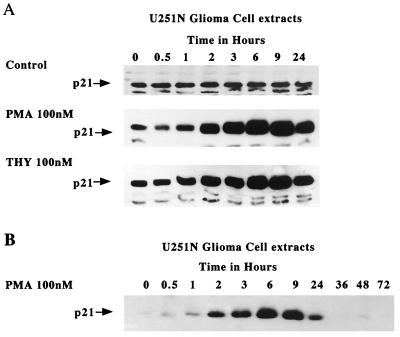
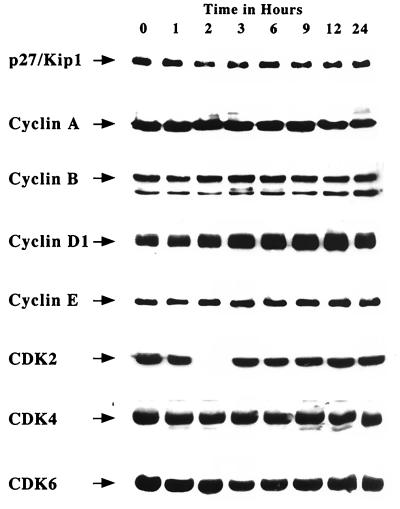
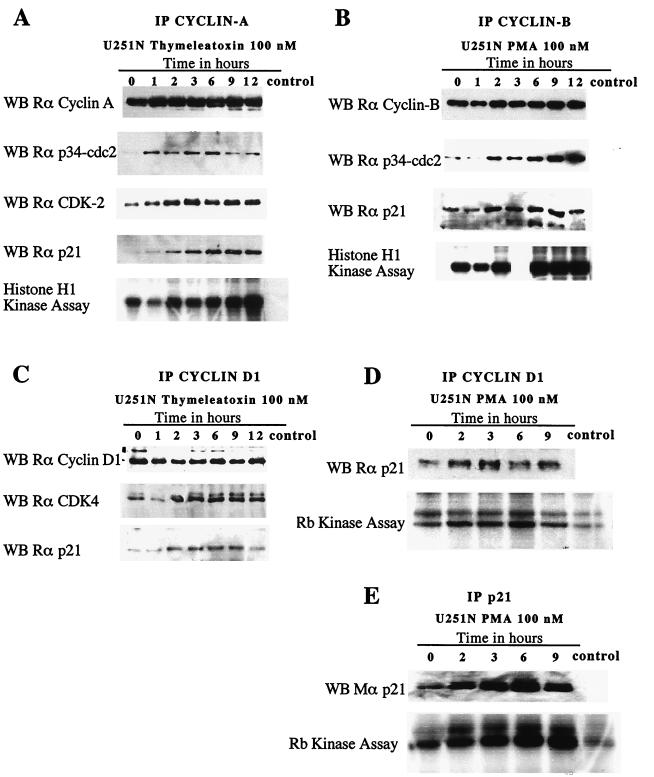
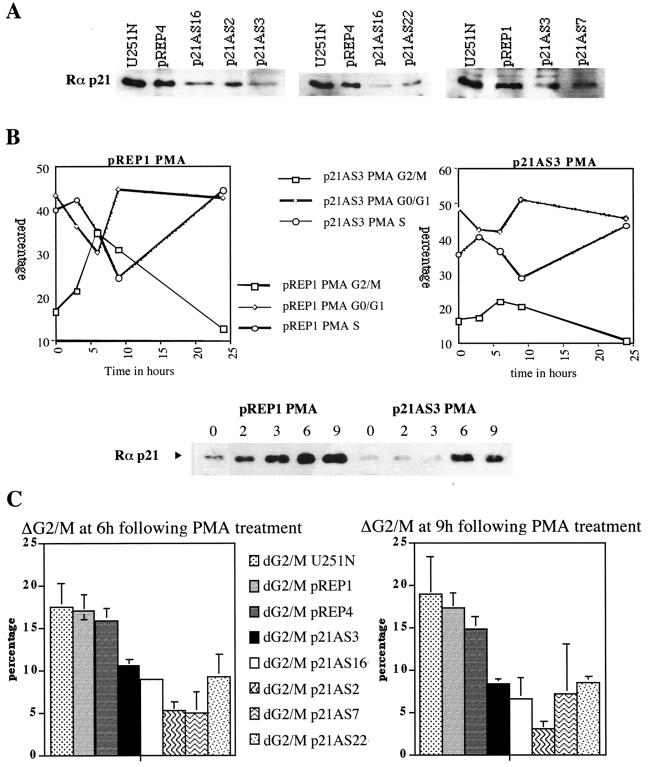
Protein kinase C (PKC) plays an important role in the regulation of glioma growth; however, the identity of the specific isoform and mechanism by which PKC fulfills this function remain unknown. In this study, we demonstrate that PKC activation in glioma cells increased their progression through the cell cycle. Of the six PKC isoforms that were present in glioma cells, PKC alpha was both necessary and sufficient to promote cell cycle progression when stimulated with phorbol 12-myristate 13-acetate. Also, decreased PKC alpha expression resulted in a marked decrease in cell proliferation. The only cell cycle-regulatory molecule whose expression was rapidly altered and increased by PKC alpha activity was the cyclin-cyclin-dependent kinase (CDK) inhibitor p21(Waf1/Cip1). Coimmunoprecipitation studies revealed that p21(Waf1/Cip1) upregulation was accompanied by an incorporation of p21(Waf1/Cip1) into various cyclin-CDK complexes and that the kinase activity of these complexes was increased, thus resulting in cell cycle progression. Furthermore, depletion of p21(Waf1/Cip1) by antisense strategy attenuated the PKC-induced cell cycle progression. These results suggest that PKC alpha activity controls glioma cell cycle progression through the upregulation of p21(Waf1/Cip1), which facilitates active cyclin-CDK complex formation.
Figures
References
-
- Ahmad S, Mineta T, Martuza R L, Glazer R I. Antisense expression of protein kinase C alpha inhibits the growth and tumorigenicity of human glioblastoma cells. Neurosurgery. 1994;35:904–909. - PubMed
-
- Arita Y, Buffolino P, Coppock D L. Regulation of the cell cycle at the G2/M boundary in metastatic melanoma cells by 12-O-tetradecanoyl phorbol-13-acetate (TPA) by blocking p34cdc2 kinase activity. Exp Cell Res. 1998;242:381–390. - PubMed
-
- Baltuch G H, Shenouda G, Langleben A, Villemure J H. High dose tamoxifen in the treatment of recurrent high-grade glioma: a report of clinical stabilization and tumor regression. Can J Neurol Sci. 1993;20:168–170. - PubMed
-
- Baltuch G H, Dooley N P, Couldwell W T, Yong V W. Staurosporine differentially inhibits glioma versus non-glioma cell lines. J Neurooncol. 1993;16:141–147. - PubMed
-
- Baltuch G H, Dooley N P, Villemure J-G, Yong V W. Protein kinase C and growth regulation of malignant gliomas. Can J Neurol Sci. 1995;22:264–271. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous