

The Ste20 kinase misshapen regulates both photoreceptor axon targeting and dorsal closure, acting downstream of distinct signals
- PMID: 10848599
- PMCID: PMC85899
- DOI: 10.1128/MCB.20.13.4736-4744.2000
The Ste20 kinase misshapen regulates both photoreceptor axon targeting and dorsal closure, acting downstream of distinct signals
Abstract
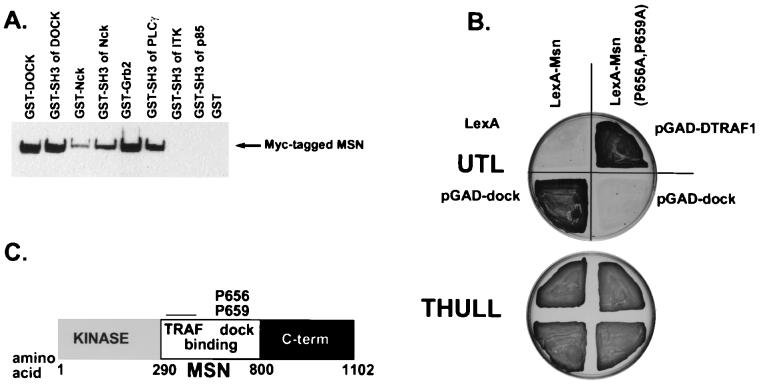
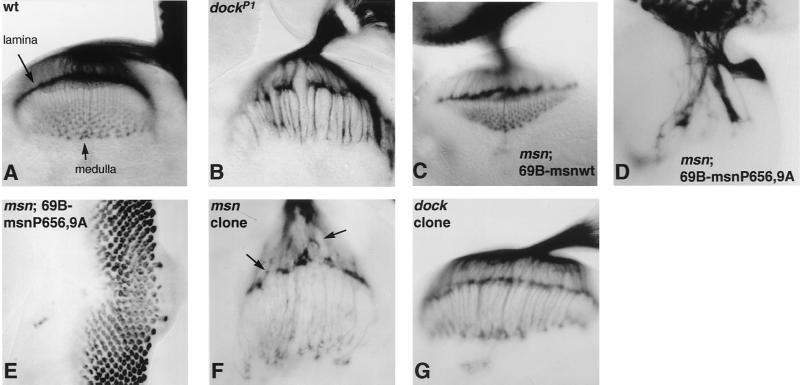
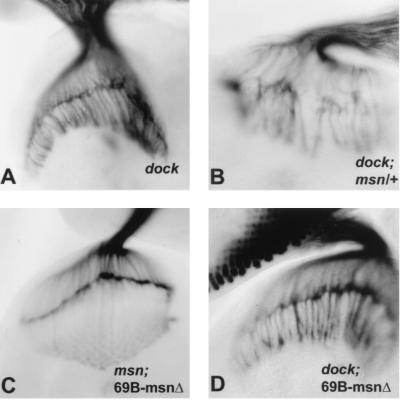
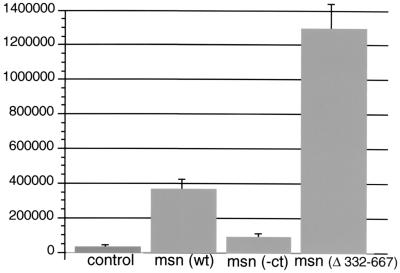
We have previously shown that the Ste20 kinase encoded by misshapen (msn) functions upstream of the c-Jun N-terminal kinase (JNK) mitogen-activated protein kinase module in Drosophila. msn is required to activate the Drosophila JNK, Basket (Bsk), to promote dorsal closure of the embryo. A mammalian homolog of Msn, Nck interacting kinase, interacts with the SH3 domains of the SH2-SH3 adapter protein Nck. We now show that Msn likewise interacts with Dreadlocks (Dock), the Drosophila homolog of Nck. dock is required for the correct targeting of photoreceptor axons. We have performed a structure-function analysis of Msn in vivo in Drosophila in order to elucidate the mechanism whereby Msn regulates JNK and to determine whether msn, like dock, is required for the correct targeting of photoreceptor axons. We show that Msn requires both a functional kinase and a C-terminal regulatory domain to activate JNK in vivo in Drosophila. A mutation in a PXXP motif on Msn that prevents it from binding to the SH3 domains of Dock does not affect its ability to rescue the dorsal closure defect in msn embryos, suggesting that Dock is not an upstream regulator of msn in dorsal closure. Larvae with only this mutated form of Msn show a marked disruption in photoreceptor axon targeting, implicating an SH3 domain protein in this process; however, an activated form of Msn is not sufficient to rescue the dock mutant phenotype. Mosaic analysis reveals that msn expression is required in photoreceptors in order for their axons to project correctly. The data presented here genetically link msn to two distinct biological events, dorsal closure and photoreceptor axon pathfinding, and thus provide the first evidence that Ste20 kinases of the germinal center kinase family play a role in axonal pathfinding. The ability of Msn to interact with distinct classes of adapter molecules in dorsal closure and photoreceptor axon pathfinding may provide the flexibility that allows it to link to distinct upstream signaling systems.
Figures
References
-
- Affolter M, Nellen D, Nussbaumer U, Basler K. Multiple requirements for the receptor serine/threonine kinase thick veins reveal novel functions of TGFβ homologs during Drosophila embryogenesis. Development. 1994;120:3105–3117. - PubMed
-
- Aronheim A, Engelberg D, Li N, al-Alawi N, Schlessinger J, Karin M. Membrane targeting of the nucleotide exchange factor Sos is sufficient for activating the Ras signaling pathway. Cell. 1994;78:949–961. - PubMed
-
- Brand A H, Perrimon N. Targeting gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Brummel T J, Twombly G, Marques G, Wrana J L, Newfeld S J, Attisano L, Massague J. Characterization and relationship of Dpp receptors encoded by the saxophone and thick veins genes in Drosophila. Cell. 1994;78:251–261. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous