

doi: 10.1128/MCB.20.13.4773-4781.2000.
Role of DBP in the circadian oscillatory mechanism
Affiliations
- PMID: 10848603
- PMCID: PMC85912
- DOI: 10.1128/MCB.20.13.4773-4781.2000
Item in Clipboard
Role of DBP in the circadian oscillatory mechanism
Mol Cell Biol.
2000 Jul.
Abstract
Transcript levels of DBP, a member of the PAR leucine zipper transcription factor family, exhibit a robust rhythm in suprachiasmatic nuclei, the mammalian circadian center. Here we report that DBP is able to activate the promoter of a putative clock oscillating gene, mPer1, by directly binding to the mPer1 promoter. The mPer1 promoter is cooperatively activated by DBP and CLOCK-BMAL1. On the other hand, dbp transcription is activated by CLOCK-BMAL1 through E-boxes and inhibited by the mPER and mCRY proteins, as is the case for mPer1. Thus, a clock-controlled dbp gene may play an important role in central clock oscillation.
Figures
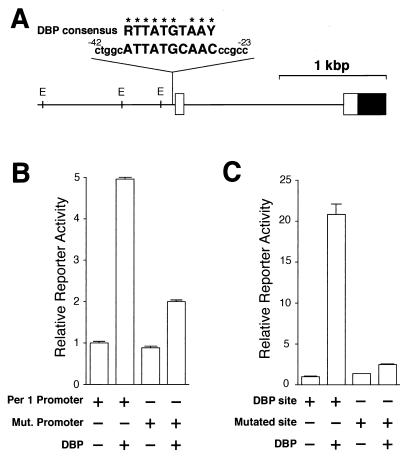
Transcriptional regulation of the mPer1 promoter by DBP. (A) A long vertical line indicates the site that matches the consensus DBP-binding sequence in the 5′-flanking region of the mPer1 gene. The sequence of that site and 5 bp of flanking sequence (lowercase letters) on each side and the consensus DBP-binding sequence (R, purine; Y, pyrimidine) are noted (identical bases are shown by asterisks). The numbers of the nucleotide residues indicate the distance from the transcription start site. The solid and open boxes show the protein-coding region and the 5′ untranslated region, respectively. The locations of the E-box sites (E) are also indicated by short vertical lines. (B) Transcriptional activation of the reporter plasmid including the mPer1 promoter. Reporter plasmids containing a 1.3-kbp fragment including the putative DBP-binding site (ATTATGCAAC) (Per1 promoter) or a mutated site (TCGCCATGGC) (Mut. promoter) were used for the transcriptional assay. (C) Transcriptional activation of an HSV-TK-driven reporter plasmid containing the putative DBP-binding site. A 60-bp construct in which three copies of either the putative DBP-binding site (DBP site) or a mutated site (TCACCCGGCT) (Mutated site) and flanking sequence were linked in tandem was subcloned into the HSV-TK-driven reporter plasmid. (B and C) Presence (+) or absence (−) of reporter and DBP expression plasmids (750 ng) is noted. Each value is the mean + SEM of three replicates for a single assay. The results shown are representative of at least three independent experiments.
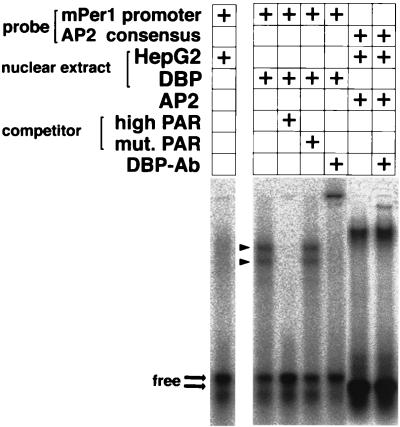
Binding of DBP to the mPer1 promoter. Gel shift assays with a probe encompassing the putative DBP-binding site derived from the mPer1 promoter were performed. Nuclear extracts from untransfected (HepG2) or DBP expression plasmid-transfected (DBP) HepG2 cells were incubated with the probe. For the competition experiment, a 100-fold excess of a double-stranded oligonucleotide containing the PAR protein recognition sequence GTTACGTAAT (high PAR) or containing the mutated sequence TCGCCATGGC (mut.PAR) was added. For the supershift experiment, 0.25 μl of DBP antiserum was added. The complex of human AP2 and an AP2 consensus binding probe was not affected by adding the DBP antiserum. The presence (+) of each probe, nuclear extract, AP2, competitor, and anti-DBP antibody is noted. Arrowheads indicate two bands representing complexes of DBP and the probe. Thick arrows point to the locations of the free probes.
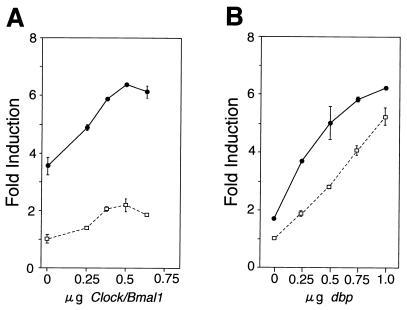
Transcriptional activation of the mPer1 promoter by DBP and CLOCK-BMAL1. (A) Dose-response curve of CLOCK-BMAL1-mediated transcriptional activation in the presence and absence of DBP. The reporter plasmid which contains the 1.3-kbp mPer1 promoter, including the three E-boxes and the DBP-responsive site, was used. Increasing doses of CLOCK and BMAL1 expression plasmids were transfected with (solid line) or without (dashed line) 750 ng of DBP expression plasmid. The total amount of DNA (2 μg) per well was adjusted by adding pcDNA3 vector. (B) Dose-response curve of DBP-mediated transcriptional activation in the presence and absence of CLOCK-BMAL1. Increasing doses of DBP expression plasmid were transfected with (solid line) or without (dashed line) 500 ng each of CLOCK and BMAL1 expression plasmids. (A and B) Each value is the mean ± SEM of duplicate for a single assay. A similar pattern of activation was reproduced in another experiment.
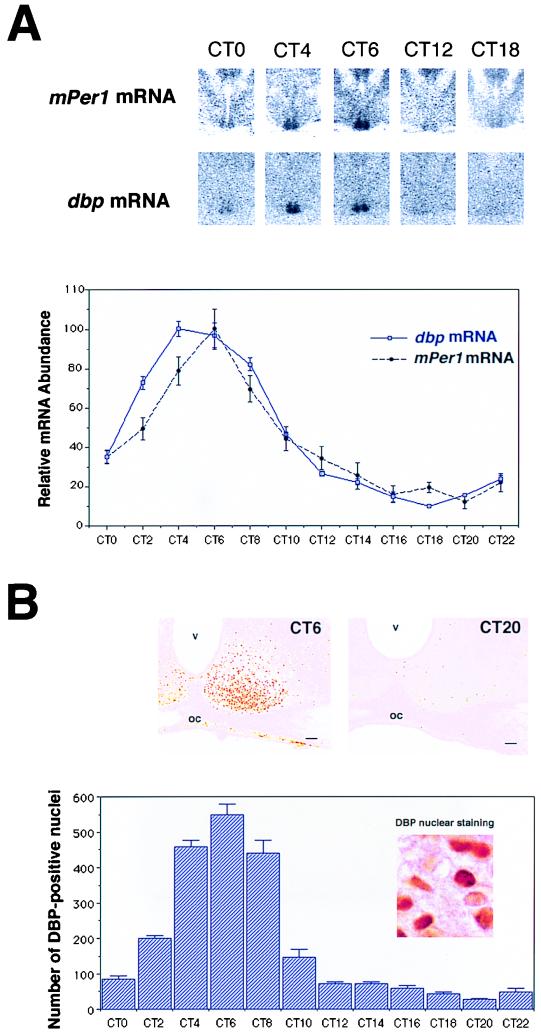
Circadian expression of dbp and mPer1 mRNA and DBP protein in the SCN. (A) Quantitative analysis of dbp (open square with blue line) and mPer1 (solid circle with dotted black line) mRNA expressed in the SCN in DD conditions (n = 5, mean ± SEM). Relative dbp and mPer1 mRNA abundance was determined by quantitative in situ hybridization using isotope-labeled probes with the mean peak values adjusted to 100. Representative in situ hybridization autoradiograms at specific time points are shown on the top panels. (B) Circadian expression of DBP immunoreactivity in the SCN. The photomicrographs show immunoreactivity at CT6 and CT20. Cell counts of SCN nuclei staining positive for DBP (per section) are shown. Values are means + SEM for four animals. oc, optic chiasma; v, third ventricle. Bar, 70 μm.
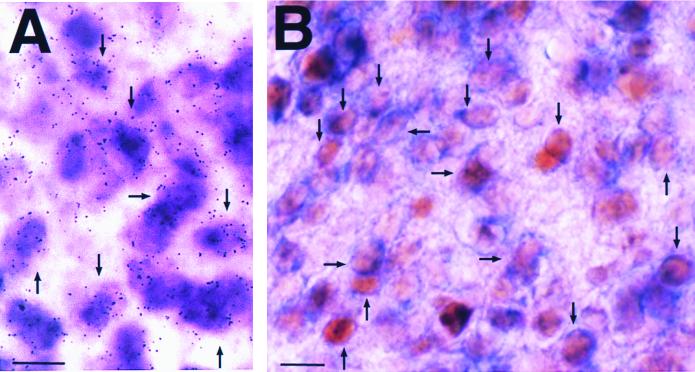
Coexpression of dbp transcripts and DBP proteins with mPer1 transcripts. (A) Double-labeling in situ hybridization using digoxigenin-labeled mPer1 probe and isotope-labeled dbp probe. Note that most cells express both mPer1 (purple) and dbp (silver grains) mRNAs. (B) Double labeling of mPer1 in situ hybridization and DBP protein immunocytochemistry. Note that the blue stain of digoxigenin-labeled mPer1 mRNA in the thin cytoplasm surrounds the brown-stained DBP protein localized in the nucleus. Arrows indicate representative double-labeled cells. All SCN sections were sampled at CT6. Bar, 10 μm.
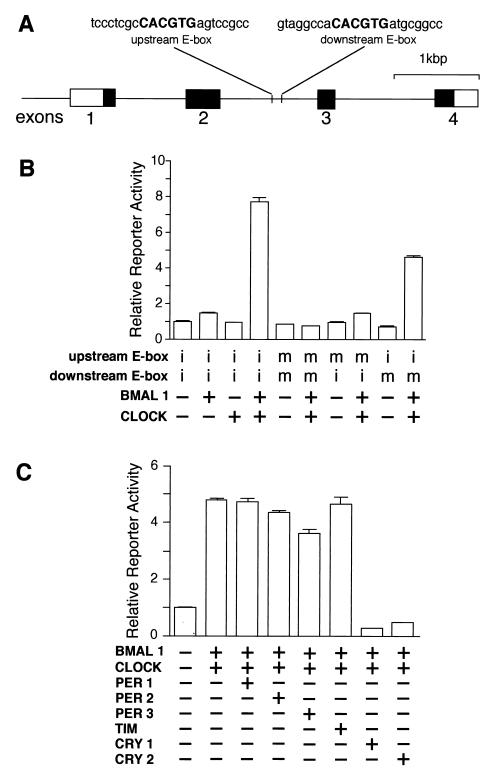
Transcriptional regulation of the mouse dbp gene by clock genes. (A) Structure of the mouse dbp gene and locations of the E-box sites. The solid and open boxes show the protein-coding region and the 5′ and 3′ untranslated regions, respectively. The locations of the E-box sites are indicated by vertical lines, and the sequence of each E-box with 8 bp of flanking sequence (lowercase letters) on each side is shown at the top. (B) Transactivation from the E-box sites by CLOCK-BMAL1 heterodimer. For transcriptional analysis, an SV40-driven reporter containing a 300-bp fragment derived from the mouse dbp gene centered on the two E-boxes was constructed and used (2.5 ng). The fragment was inserted immediately upstream of the SV40 promoter in the native 5′-to-3′ direction. Reporter plasmids in which one or two E-boxes were mutated were also constructed and used. Intact (i) or mutated (m) E-box of each reporter plasmid is shown, and 500 ng each of expression plasmids was used. (C) Negative regulation of CLOCK-BMAL1-induced transcription. The reporter plasmid (5 ng) containing the 300-bp fragment with the intact E-boxes was used. The amounts of the expression plasmids used were 750 ng for BMAL1 and CLOCK; 125 ng for PER1, PER2, PER3, and mTIM; and 50 ng for CRY1 and CRY2. The total amount (2 μg) of DNA was adjusted by adding pcDNA3 vector. (B and C) Presence (+) or absence (−) of the expression plasmids is shown. Each value is the mean + SEM of three replicates for a single assay. The results shown are representative of at least three independent experiments.
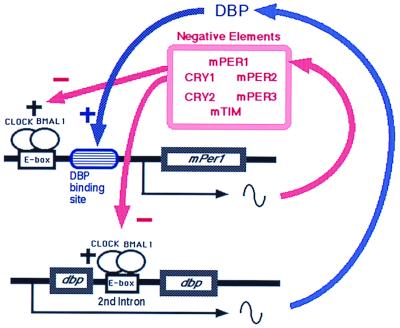
Schematic representation of the role of DBP in the circadian oscillatory mechanism of the SCN. In addition to the negative autoregulatory feedback loop of mPer1, a DBP-mediated loop exists. As DBP protein positively regulates the mPer1 promoter, DBP amplifies the circadian oscillation of mPer1 and thereby influences the circadian oscillator.
References
-
- Albrecht U, Sun Z S, Eichele G, Lee C C. A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light. Cell. 1997;91:1055–1064. - PubMed
-
- Blau J, Young M W. Cycling vrille expression is required for a functional Drosophila clock. Cell. 1999;99:661–671. - PubMed
-
- Citri Y, Colot H V, Jacquier A C, Yu Q, Hall J C, Baltimore D, Rosbach M. A family of unusually spliced biologically active transcripts encoded by a Drosophila clock gene. Nature. 1987;326:42–47. - PubMed
-
- Dunlap J C. Molecular bases for circadian clocks. Cell. 1999;96:271–290. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources