

Spatially regulated recruitment of clathrin to the plasma membrane during capping and cell translocation
- PMID: 10848635
- PMCID: PMC14909
- DOI: 10.1091/mbc.11.6.2151
Spatially regulated recruitment of clathrin to the plasma membrane during capping and cell translocation
Abstract
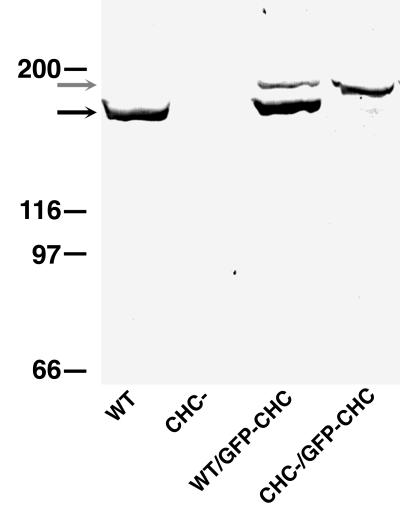
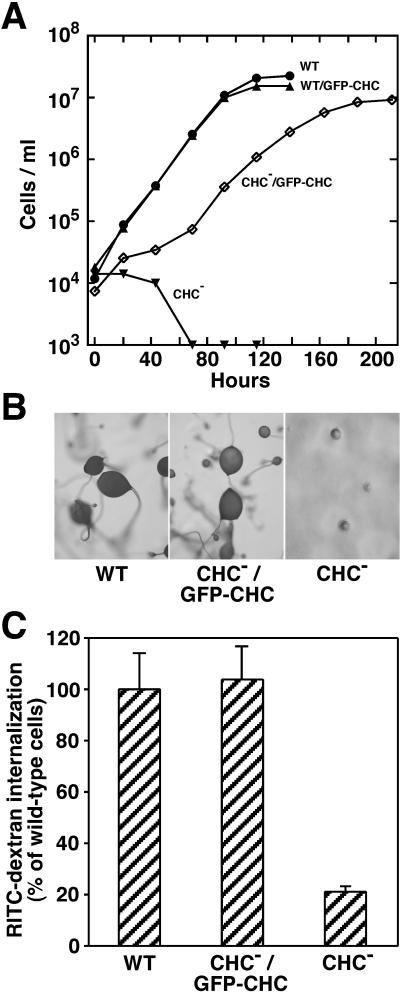
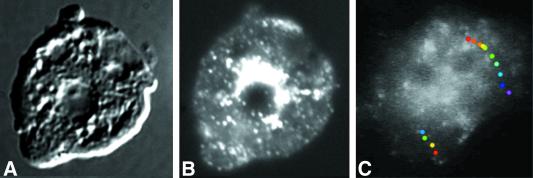
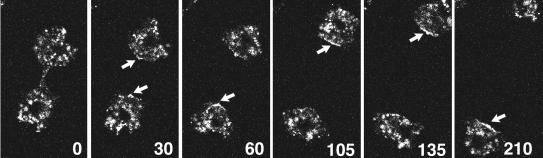
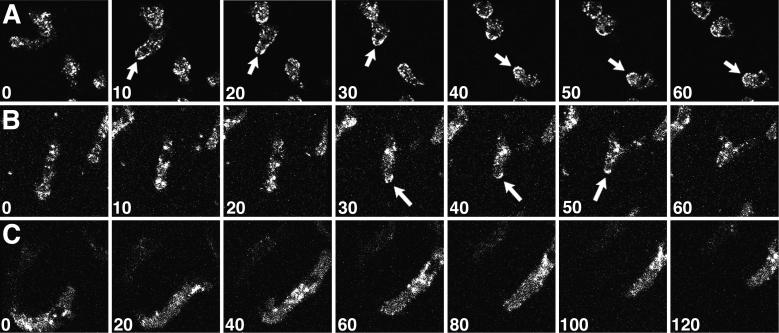
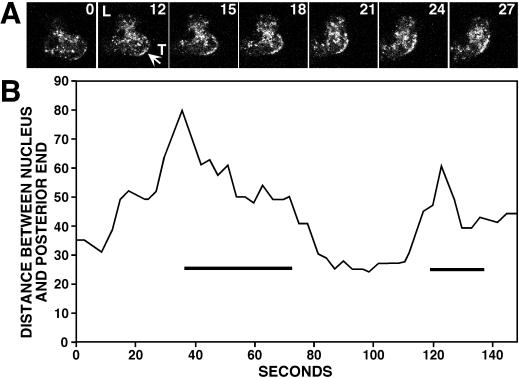
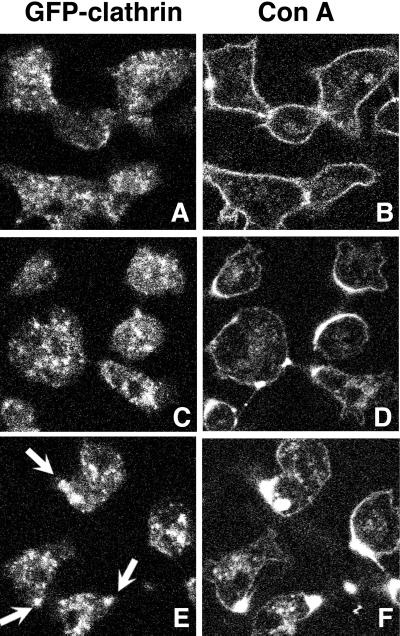
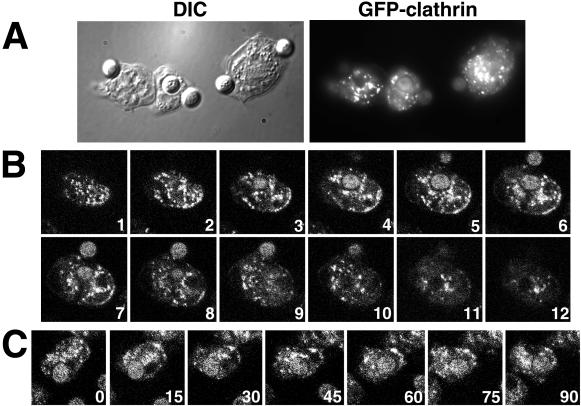
Clathrin-coated vesicles bud from selected cellular membranes to traffic-specific intracellular proteins. To study the dynamic properties of clathrin-coated membranes, we expressed clathrin heavy chain tagged with green fluorescent protein (GFP) in Dictyostelium cells. GFP-clathrin was functional and retained the native properties of clathrin: the chimeric protein formed classic clathrin lattices on cellular membranes and also rescued phenotypic defects of clathrin null cells. GFP-clathrin distributed into punctate loci found throughout the cytoplasm, on the plasma membrane, and concentrated to a perinuclear location. These clathrin-coated structures were remarkably motile and capable of rapid and bidirectional transport across the cell. We identified two local domains of the plasma membrane as sites for clathrin recruitment in motile cells. First, as cells translocated or changed shape and retracted their tails, clathrin was transiently concentrated on the membrane at the back of the cell tail. Second, as cells capped their cell surface receptors, clathrin was recruited locally to the membrane under the tight cap of cross-linked receptors. This suggests that local sites for clathrin polymerization on specific domains of the plasma membrane undergo rapid and dynamic regulation in motile cells.
Figures
References
-
- Anderson RGW, Brown MS, Goldstein JL. Role of the coated endocytic vesicle in the uptake of receptor-bound low density lipoprotein in human fibroblasts. Cell. 1977;10:351–364. - PubMed
-
- Bretscher MS. Endocytosis: relation to capping and cell locomotion. Science. 1984;224:681–686. - PubMed
-
- Bretscher MS. How animal cells move. Sci Am. 1987;257:72–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
