

The tyrosine kinase PYK-2/RAFTK regulates natural killer (NK) cell cytotoxic response, and is translocated and activated upon specific target cell recognition and killing
- PMID: 10851022
- PMCID: PMC2175114
- DOI: 10.1083/jcb.149.6.1249
The tyrosine kinase PYK-2/RAFTK regulates natural killer (NK) cell cytotoxic response, and is translocated and activated upon specific target cell recognition and killing
Abstract
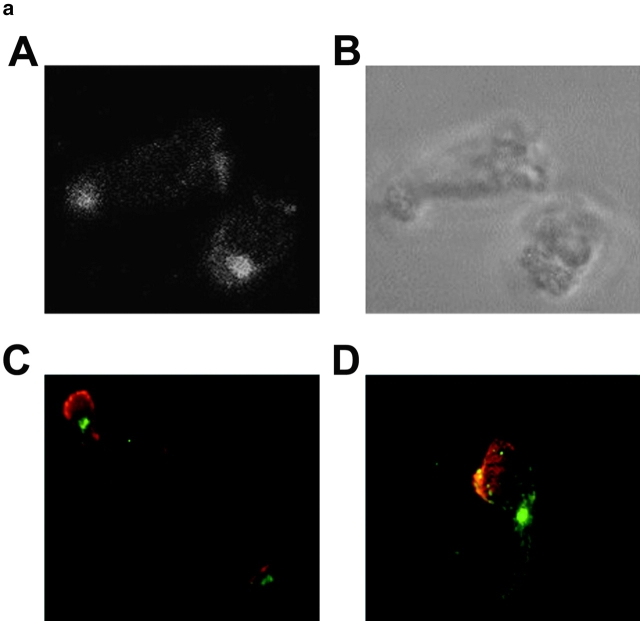
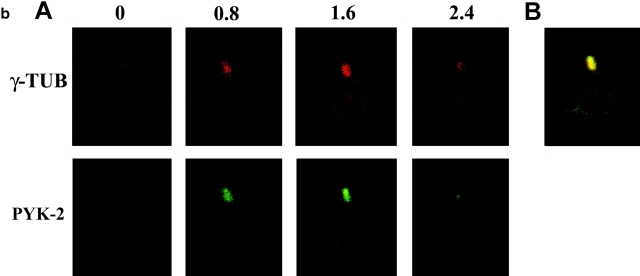
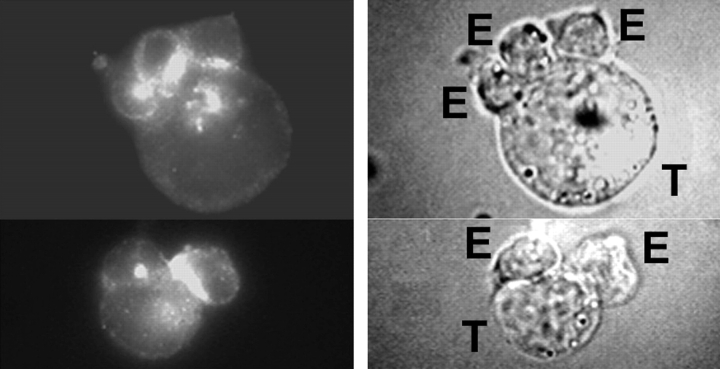
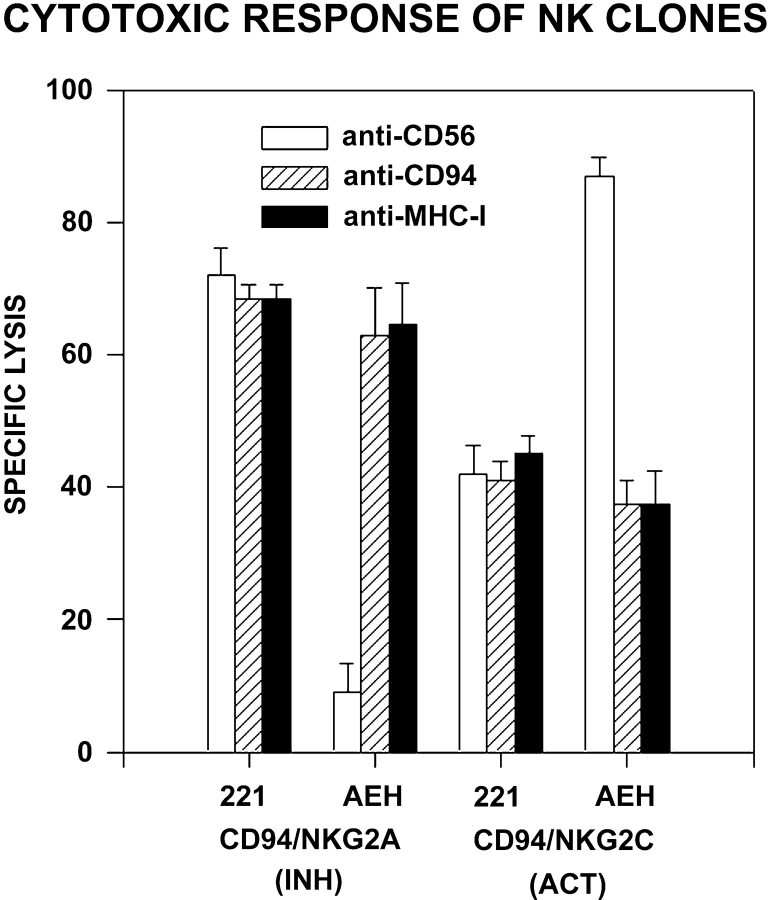
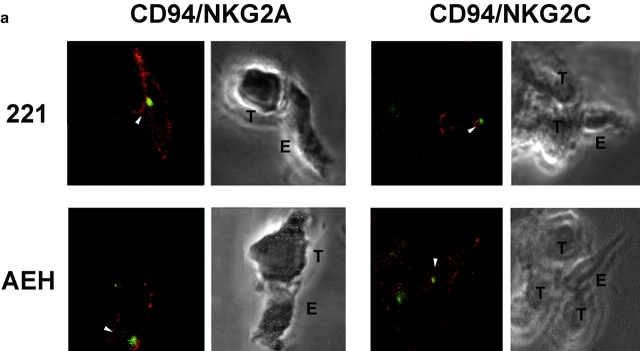
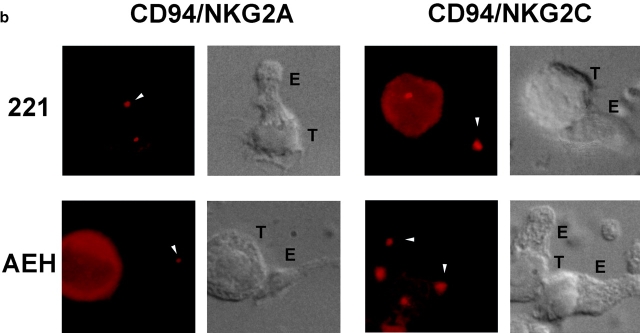
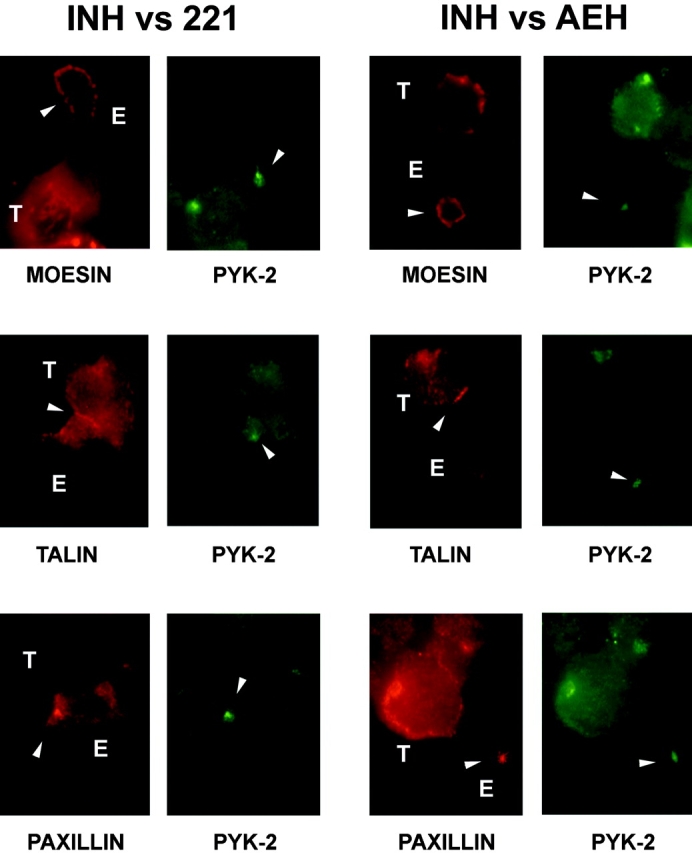
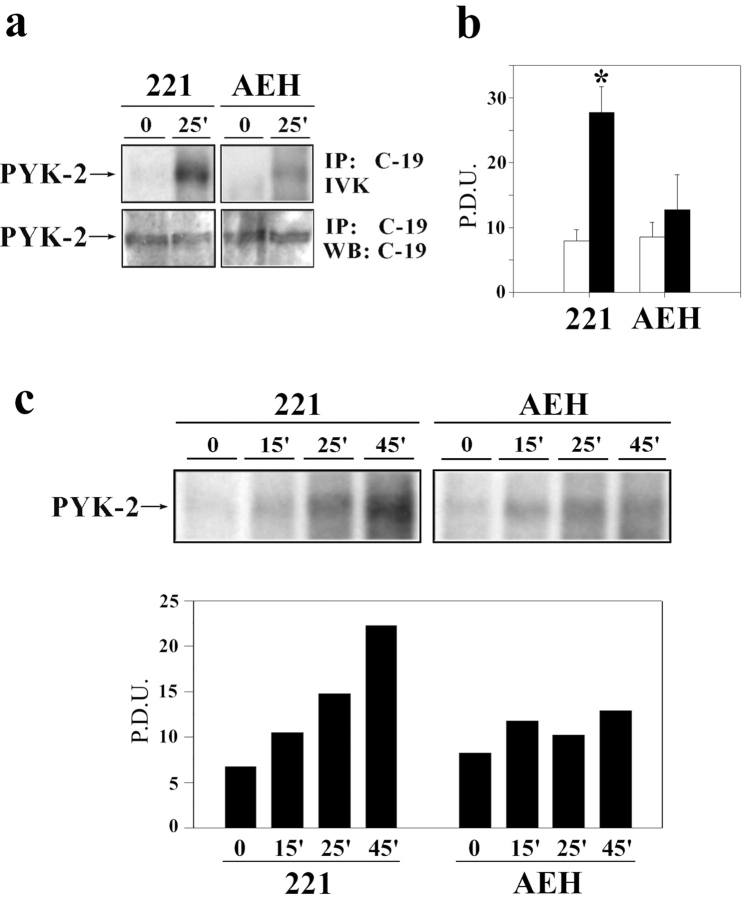
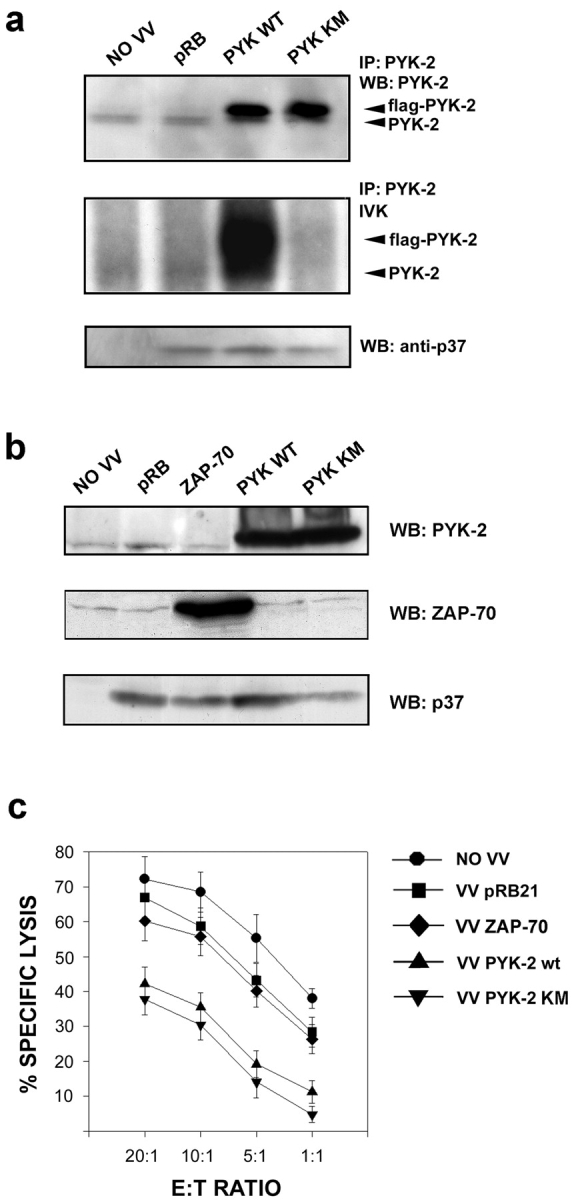
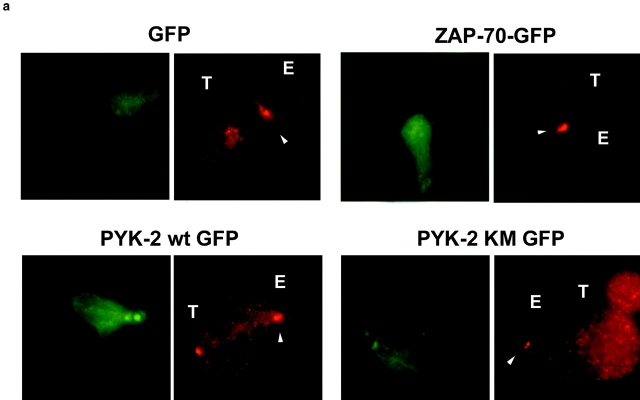
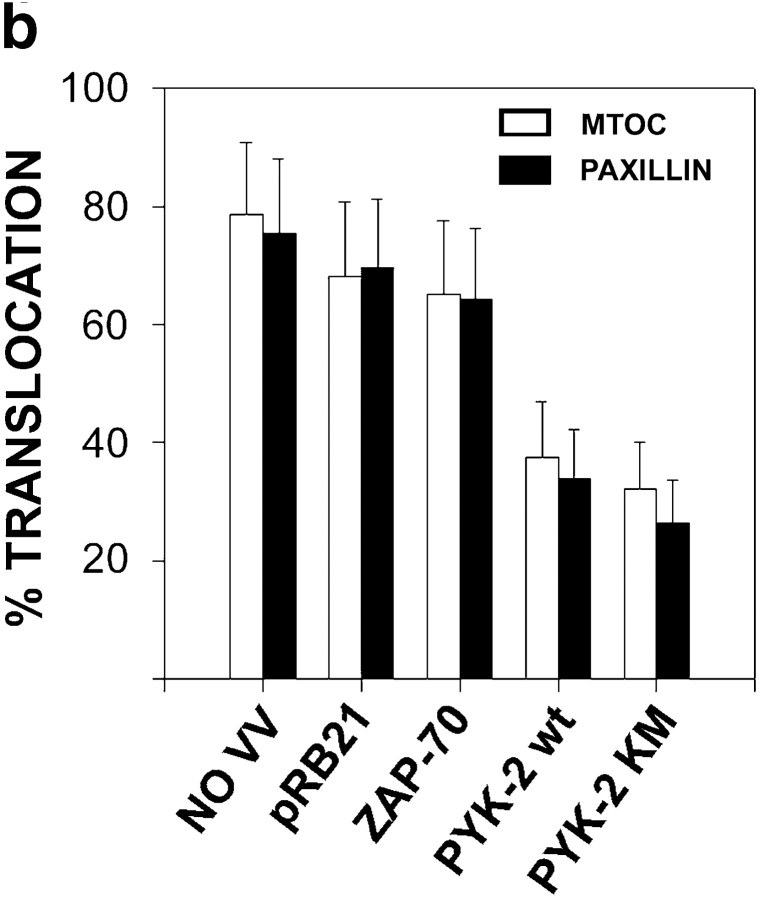
The compartmentalization of plasma membrane proteins has a key role in regulation of lymphocyte activation and development of immunity. We found that the proline-rich tyrosine kinase-2 (PYK-2/RAFTK) colocalized with the microtubule-organizing center (MTOC) at the trailing edge of migrating natural killer (NK) cells. When polyclonal NK cells bound to K562 targets, PYK-2 translocated to the area of NK-target cell interaction. The specificity of this process was assessed with NK cell clones bearing activatory or inhibitory forms of CD94/NKG2. The translocation of PYK-2, MTOC, and paxillin to the area of NK-target cell contact was regulated upon specific recognition of target cells through NK cell receptors, controlling target cell killing. Furthermore, parallel in vitro kinase assays showed that PYK-2 was activated in response to signals that specifically triggered its translocation and NK cell mediated cytotoxicity. The overexpression of both the wt and a dominant-negative mutant of PYK-2, but not ZAP-70 wt, prevented the specific translocation of the MTOC and paxillin, and blocked the cytotoxic response of NK cells. Our data indicate that subcellular compartmentalization of PYK-2 correlates with effective signal transduction. Furthermore, they also suggest an important role for PYK-2 on the assembly of the signaling complexes that regulate the cytotoxic response.
Figures
Similar articles
-
Proline-rich tyrosine kinase-2 activation by beta 1 integrin fibronectin receptor cross-linking and association with paxillin in human natural killer cells.J Immunol. 1997 Nov 15;159(10):4729-36. J Immunol. 1997. PMID: 9366396
-
CD94/NKG2-A inhibitory complex blocks CD16-triggered Syk and extracellular regulated kinase activation, leading to cytotoxic function of human NK cells.J Immunol. 1999 Jun 15;162(12):7181-8. J Immunol. 1999. PMID: 10358164
-
Clonotypic differences in signaling from CD94 (kp43) on NK cells lead to divergent cellular responses.J Immunol. 1996 Oct 1;157(7):2804-12. J Immunol. 1996. PMID: 8816383
-
NK cell receptors.Annu Rev Immunol. 1998;16:359-93. doi: 10.1146/annurev.immunol.16.1.359. Annu Rev Immunol. 1998. PMID: 9597134 Review.
-
Regulation of the natural killer cell receptor repertoire.Annu Rev Immunol. 2001;19:291-330. doi: 10.1146/annurev.immunol.19.1.291. Annu Rev Immunol. 2001. PMID: 11244039 Review.
Cited by
-
The killer's kiss: the many functions of NK cell immunological synapses.Curr Opin Cell Biol. 2008 Oct;20(5):597-605. doi: 10.1016/j.ceb.2008.05.006. Epub 2008 Jul 17. Curr Opin Cell Biol. 2008. PMID: 18639449 Free PMC article. Review.
-
Navigating barriers: the challenge of directed secretion at the natural killer cell lytic immunological synapse.J Clin Immunol. 2010 May;30(3):358-63. doi: 10.1007/s10875-010-9372-y. Epub 2010 Feb 27. J Clin Immunol. 2010. PMID: 20191315 Free PMC article. Review.
-
CD56 on intratumoral NK cells: orchestrating NK cell-mediated anti-tumor effects in bladder cancer.Neoplasia. 2025 Aug;66:101187. doi: 10.1016/j.neo.2025.101187. Epub 2025 May 28. Neoplasia. 2025. PMID: 40441067 Free PMC article.
-
Micro-adhesion rings surrounding TCR microclusters are essential for T cell activation.J Exp Med. 2016 Jul 25;213(8):1609-25. doi: 10.1084/jem.20151088. Epub 2016 Jun 27. J Exp Med. 2016. PMID: 27354546 Free PMC article.
-
Deficient natural killer cell cytotoxicity in patients with IKK-gamma/NEMO mutations.J Clin Invest. 2002 Jun;109(11):1501-9. doi: 10.1172/JCI14858. J Clin Invest. 2002. PMID: 12045264 Free PMC article.
References
-
- Aramburu J., Balboa M.A., Ramírez A., Silva A., Acevedo A., Sánchez-Madrid F., de Landázuri M.O., López-Botet M. A novel functional cell surface dimer (Kp43) expressed by natural killer cells and T cell receptor-γδ T lymphocytes. Inhibition of IL-2 dependent proliferation by anti-Kp43 monoclonal antibody. J. Immunol. 1990;147:714–721. - PubMed
-
- Astier A., Avraham H., Manie S.N., Groopman J., Canty T., Avraham S., Freedman A.S. The related adhesion focal tyrosine kinase is tyrosine phosphorylated after β1-integrin stimulation in B cells and binds to p130cas . J. Biol. Chem. 1997;272:228–232. - PubMed
-
- Avraham S., London R., Fu Y., Ota S., Hiregowdara D., Li S., Jiang S., Pasztor L.M., White R.A., Groopman J.E., Avraham H. Identification and characterization of a novel related adhesion focal tyrosine kinase (RAFTK) from megakaryocytes and brain. J. Biol. Chem. 1995;270:27742–27751. - PubMed
-
- Berg N.N., Ostergaard H.L. T cell receptor engagement induces tyrosine phosphorylation of FAK and PYK-2 and their association with Lck. J. Immunol. 1997;159:1753–1757. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
