

A novel shuttling protein, 4E-T, mediates the nuclear import of the mRNA 5' cap-binding protein, eIF4E
- PMID: 10856257
- PMCID: PMC203362
- DOI: 10.1093/emboj/19.12.3142
A novel shuttling protein, 4E-T, mediates the nuclear import of the mRNA 5' cap-binding protein, eIF4E
Abstract
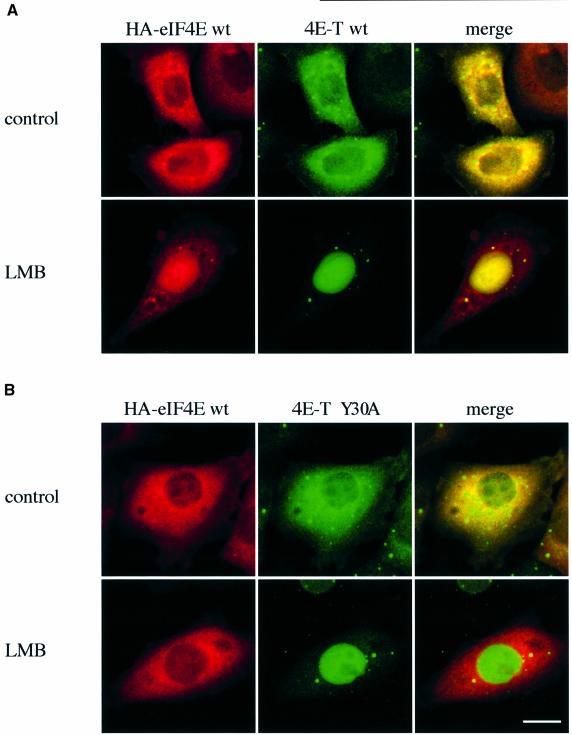
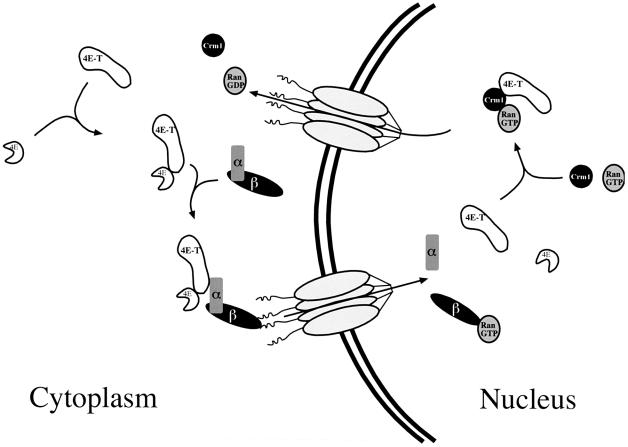
The eukaryotic translation initiation factor 4E (eIF4E) plays an important role in the control of cell growth. eIF4E binds to the mRNA 5' cap structure m(7)GpppN (where N is any nucleotide), and promotes ribosome binding to the mRNA in the cytoplasm. However, a fraction of eIF4E localizes to the nucleus. Here we describe the cloning and functional characterization of a new eIF4E-binding protein, referred to as 4E-T (eIF4E-Transporter). We demonstrate that 4E-T is a nucleocytoplasmic shuttling protein that contains an eIF4E-binding site, one bipartite nuclear localization signal and two leucine-rich nuclear export signals. eIF4E forms a complex with the importin alphabeta heterodimer only in the presence of 4E-T. Overexpression of wild-type 4E-T, but not of a mutant defective for eIF4E binding, causes the nuclear accumulation of HA-eIF4E in cells treated with leptomycin B. Taken together, these results demonstrate that the novel nucleocytoplasmic shuttling protein 4E-T mediates the nuclear import of eIF4E via the importin alphabeta pathway by a piggy-back mechanism.
Figures




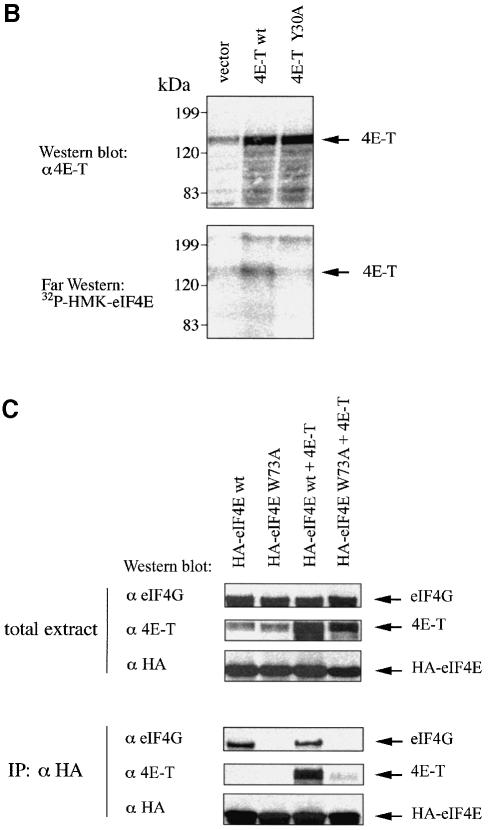
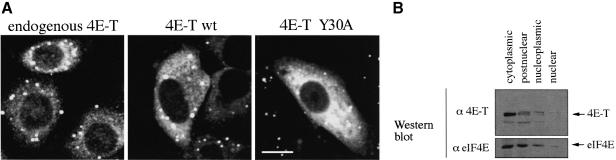
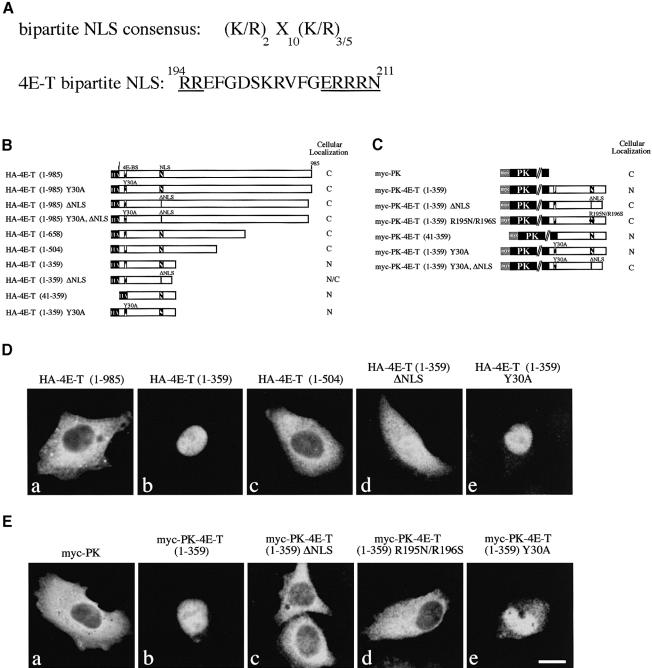
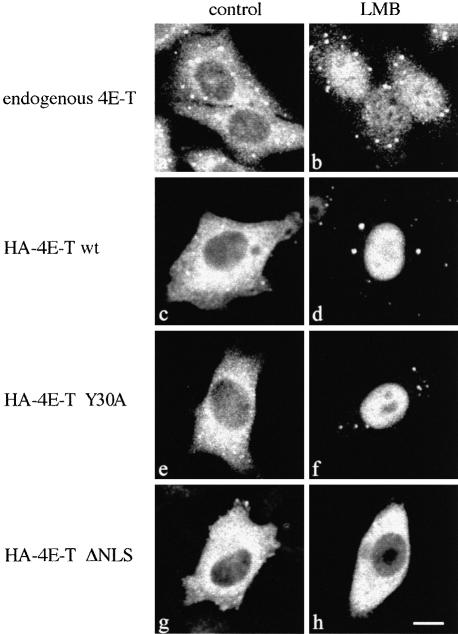


References
-
- Blanar M.A. and Rutter,W.J. (1992) Interaction cloning: identification of a helix–loop–helix zipper protein that interacts with c-Fos. Science, 256, 1014–1018. - PubMed
-
- Duncan R., Milburn,S.C. and Hershey,J.W. (1987) Regulated phosphorylation and low abundance of HeLa cell initiation factor eIF-4F suggest a role in translational control. Heat shock effects on eIF-4F. J. Biol. Chem., 262, 380–388. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
