

Full capacity of recombinant Escherichia coli heat-stable enterotoxin fusion proteins for extracellular secretion, antigenicity, disulfide bond formation, and activity
- PMID: 10858223
- PMCID: PMC101696
- DOI: 10.1128/IAI.68.7.4064-4074.2000
Full capacity of recombinant Escherichia coli heat-stable enterotoxin fusion proteins for extracellular secretion, antigenicity, disulfide bond formation, and activity
Abstract
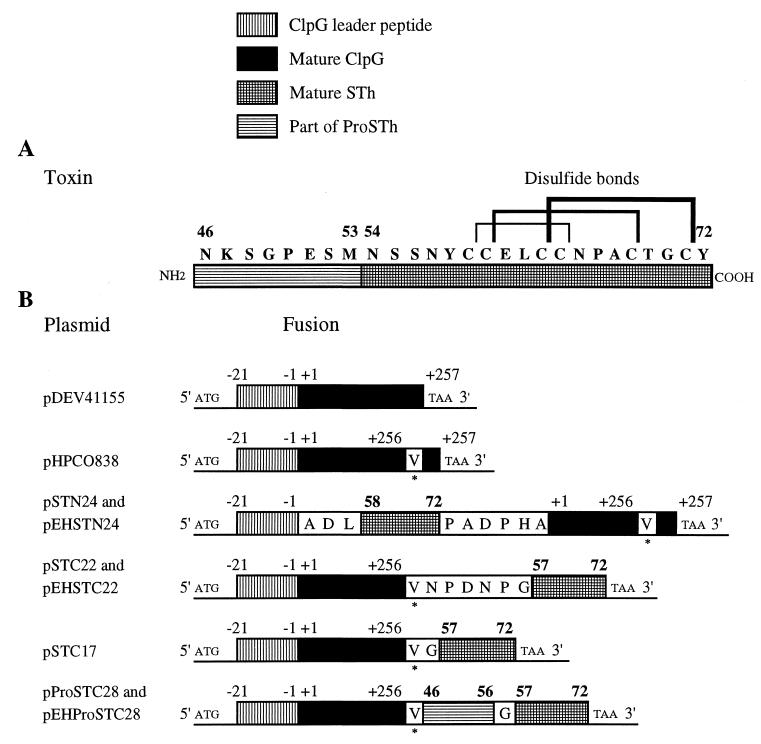
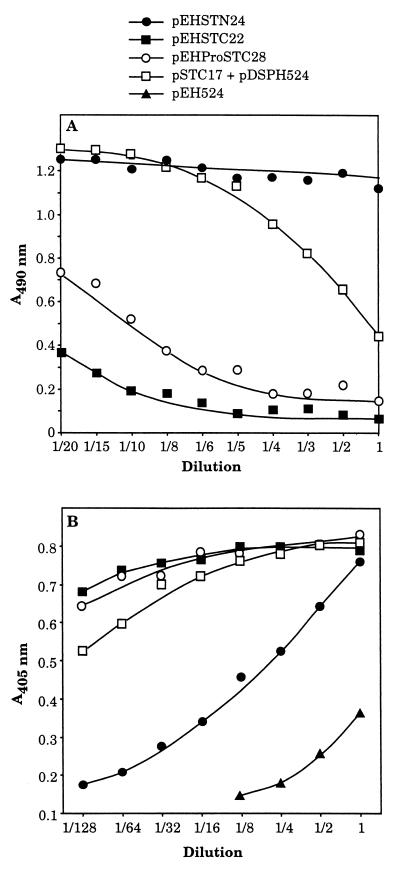
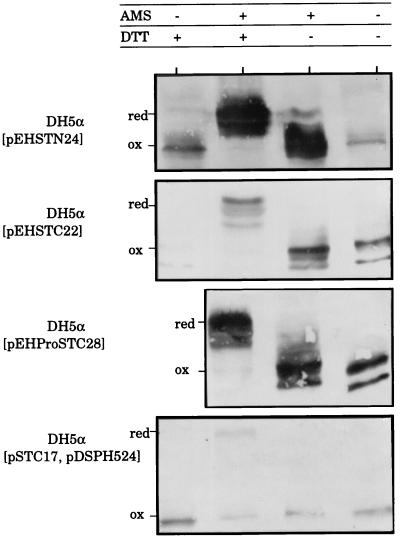
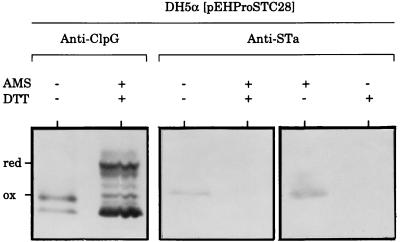
We have successfully used the major subunit ClpG of Escherichia coli CS31A fimbriae as an antigenic and immunogenic exposure-delivery vector for various heterologous peptides varying in nature and length. However, the ability of ClpG as a carrier to maintain in vitro and in vivo the native biological properties of passenger peptide has not yet been reported. To address this possibility, we genetically fused peptides containing all or part of the E. coli human heat-stable enterotoxin (STh) sequence to the amino or carboxyl ends of ClpG. Using antibodies to the ClpG and STh portions for detecting the hybrids; AMS (4-acetamido-4'-maleimidylstilbene-2, 2'-disulfonate), a potent free thiol-trapping reagent, for determining the redox state of STh in the fusion; and the suckling mouse assay for enterotoxicity, we demonstrated that all ClpG-STh proteins were secreted in vitro and in vivo outside the E. coli cells in a heat-stable active oxidized (disulfide-bonded) form. Indeed, in contrast to many earlier studies, blocking the natural NH(2) or COOH extremities of STh had, in all cases, no drastic effect on cell release and toxin activity. Only antigenicity of STh C-terminally extended with ClpG was strongly affected in a conformation-dependent manner. These results suggest that the STh activity was not altered by the chimeric structure, and therefore that, like the natural toxin, STh in the fusion had a spatial structure flexible enough to be compatible with secretion and enterotoxicity (folding and STh receptor recognition). Our study also indicates that disulfide bonds were essential for enterotoxicity but not for release, that spontaneous oxidation by molecular oxygen occurred in vitro in the medium, and that the E. coli cell-bound toxin activity in vivo resulted from an effective export processing of hybrids and not cell lysis. None of the ClpG-STh subunits formed hybrid CS31A-STh fimbriae at the cell surface of E. coli, and a strong decrease in the toxin activity was observed in the absence of CS31A helper proteins. In fact, chimeras translocated across the outer membrane as a free folded monomer once they were guided into the periplasm by the ClpG leader peptide through the CS31A-dependent secretory pathway. In summary, ClpG appears highly attractive as a carrier reporter protein for basic and applied research through the engineering of E. coli for culture supernatant delivery of an active cysteine-containing protein, such as the heat-stable enterotoxin.
Figures
References
-
- Acheson D W K. Enterotoxins in acute infective diarrhoea. J Infect. 1992;24:225–245. - PubMed
-
- Aitken R, Hirst T R. Recombinant enterotoxins as vaccines against Escherichia coli-mediated diarrhoea. Vaccine. 1993;11:227–233. - PubMed
-
- Bakker D, Vader C E M, Roosendaal B, Mooi F R, Oudega B, de Graaf F K. Structure and function of periplasmic chaperone-like proteins involved in the biosynthesis of K88 and K99 fimbriae in enterotoxigenic Escherichia coli. Mol Microbiol. 1991;5:875–886. - PubMed
-
- Bertin Y, Girardeau J P, Der Vartanian M, Martin C. The ClpE protein involved in biogenesis of the CS31A capsule-like antigen is a member of a periplasmic chaperone family in Gram-negative bacteria. FEMS Microbiol Lett. 1993;108:59–68. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
