

Effects of estradiol and progesterone on susceptibility and early immune responses to Chlamydia trachomatis infection in the female reproductive tract
- PMID: 10858238
- PMCID: PMC101727
- DOI: 10.1128/IAI.68.7.4207-4216.2000
Effects of estradiol and progesterone on susceptibility and early immune responses to Chlamydia trachomatis infection in the female reproductive tract
Abstract
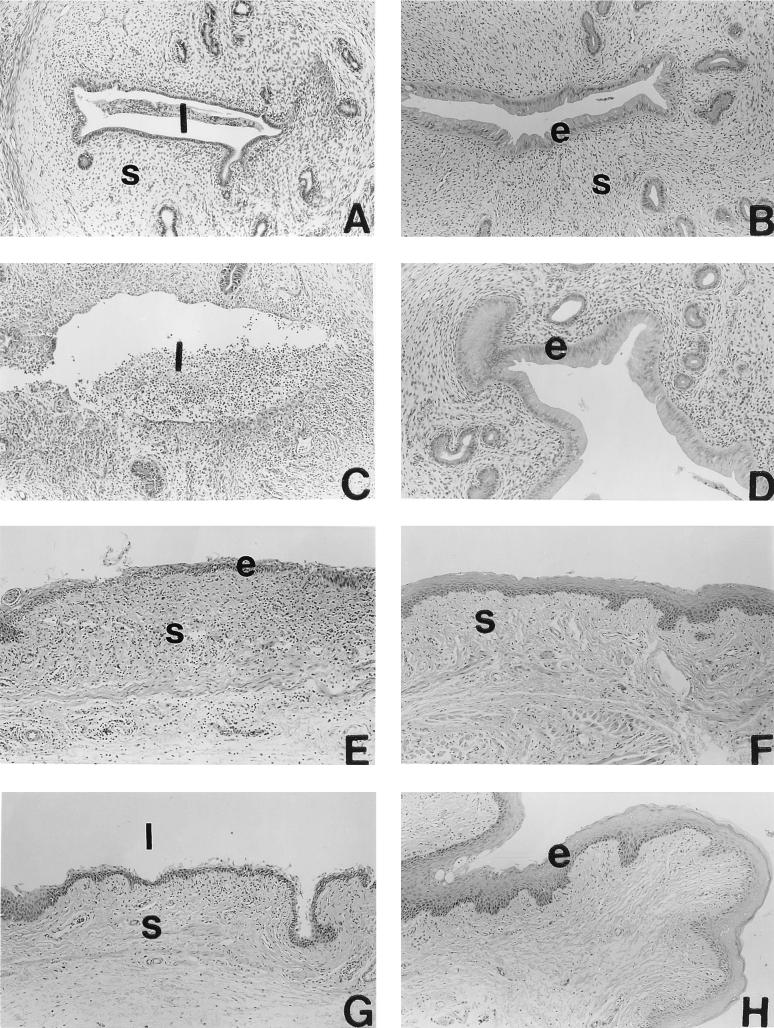
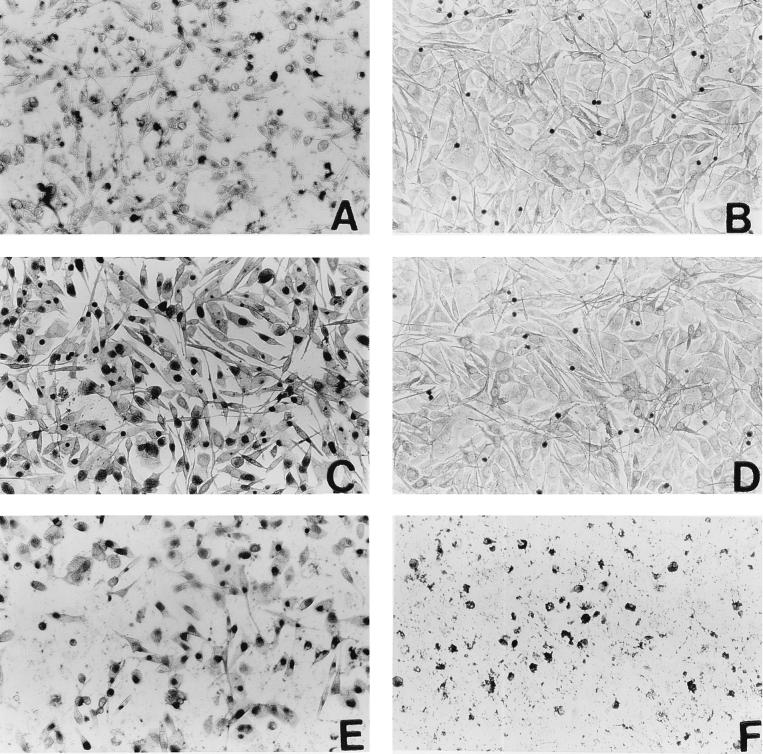
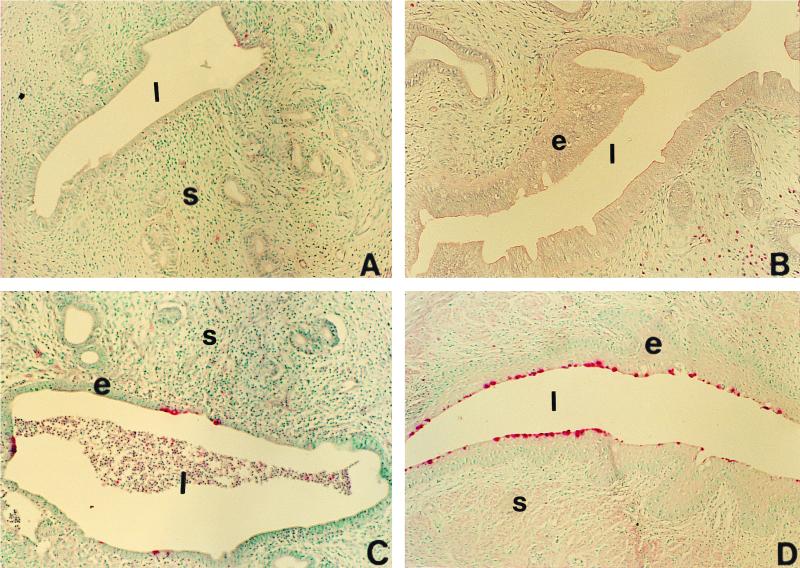
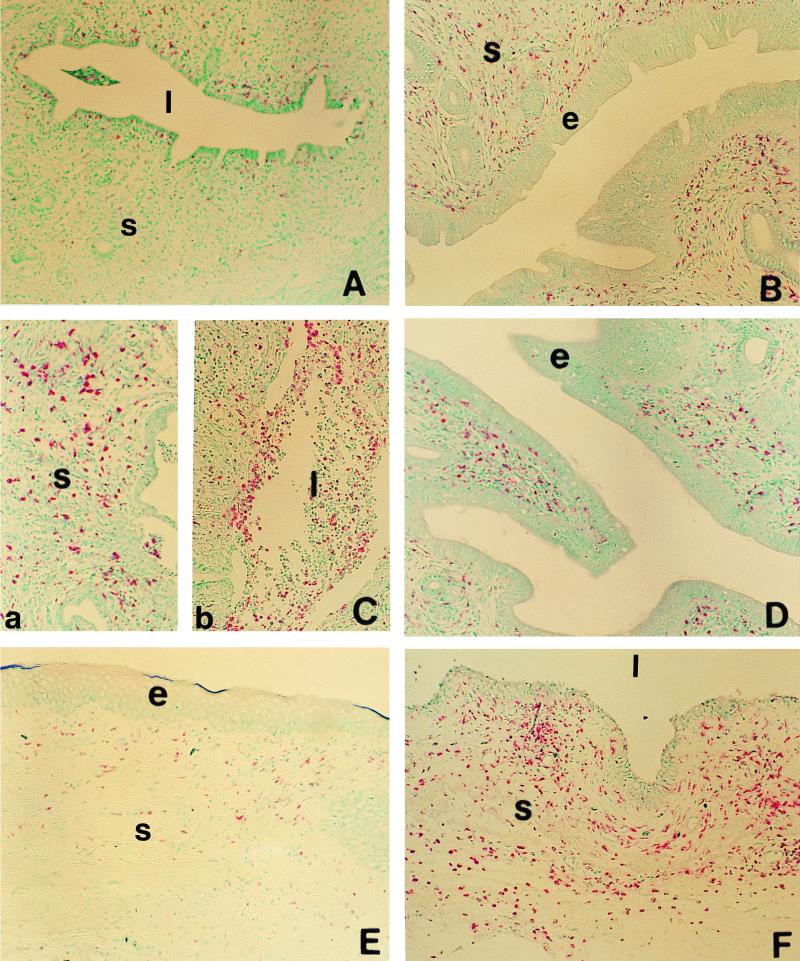
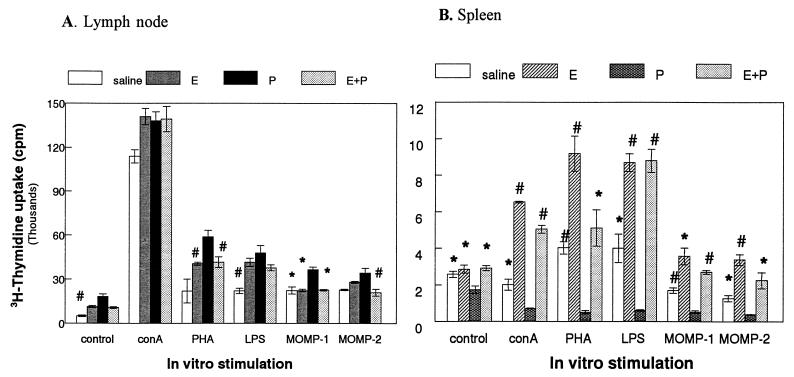
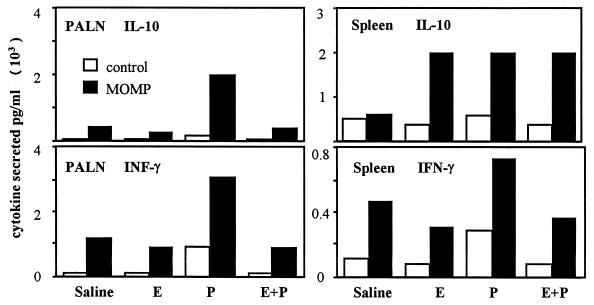
We have used a previously described rodent model to examine the influence of hormonal environment on susceptibility and immune responses to genital Chlamydia infection. Ovariectomized rats were administered estradiol, progesterone, or a combination of both, infected with Chlamydia trachomatis via the intrauterine route, and sacrificed 5 days later. Histopathological examination showed severe inflammation in the uteri and vaginae of progesterone-treated animals, whereas animals receiving estradiol or a combination of both hormones showed no inflammation. Large numbers of chlamydiae were found in vaginal secretions of progesterone-treated and combination-treated animals, while estradiol-treated animals had none. Tissue localization showed that numerous chlamydial inclusions were present in the uterine epithelium of the progesterone group and the cervicovaginal epithelium of the combination group. Examination of the acute immune responses of the infected animals showed that maximum activation was present in the draining lymph node cells from the progesterone-treated group, and these cells were producing large amounts of interleukin-10 and gamma interferon compared to other hormone-treated groups. In contrast, spleen cell proliferation was suppressed in progesterone-treated animals compared to other hormone-treated groups. We conclude that progesterone increases and estradiol decreases susceptibility to intrauterine chlamydial infection in this rat model. Our data demonstrate that hormone environment, at the time of infection, has a profound effect on the outcome of microbial infection in the female reproductive tract.
Figures
References
-
- Baker D A, Plotkin S A. Enhancement of vaginal infection in mice by herpes simplex virus type II with progesterone. Proc Soc Exp Biol Med. 1978;158:131–134. - PubMed
-
- Barron A, Pasley J N, Rank R G, White H J, Mrak R E. Chlamydial salpingitis in female guinea pigs receiving oral contraceptives. Sex Transm Dis. 1988;15:169–173. - PubMed
-
- Ito J J I, Harrison R, Alexander E R, Billings L J. Establishment of genital tract infection in the CF-1 mouse by intravaginal inoculation of a human oculogenital isolate of chlamydia trachomatis. J Infect Dis. 1984;150:577–582. - PubMed
-
- Kaushic C, Frauendorf E, Rossoll R M, Richardson J M, Wira C R. Influence of estrous cycle on the presence and distribution of immune cells in the rat reproductive tract. Am J Reprod Immunol. 1998;39:209–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
