

daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans
- PMID: 10859169
- PMCID: PMC316684
daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans
Abstract
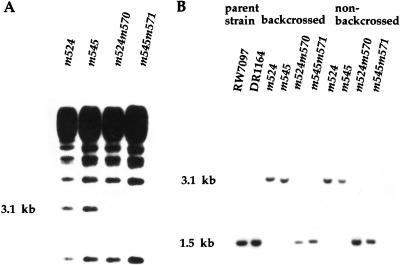
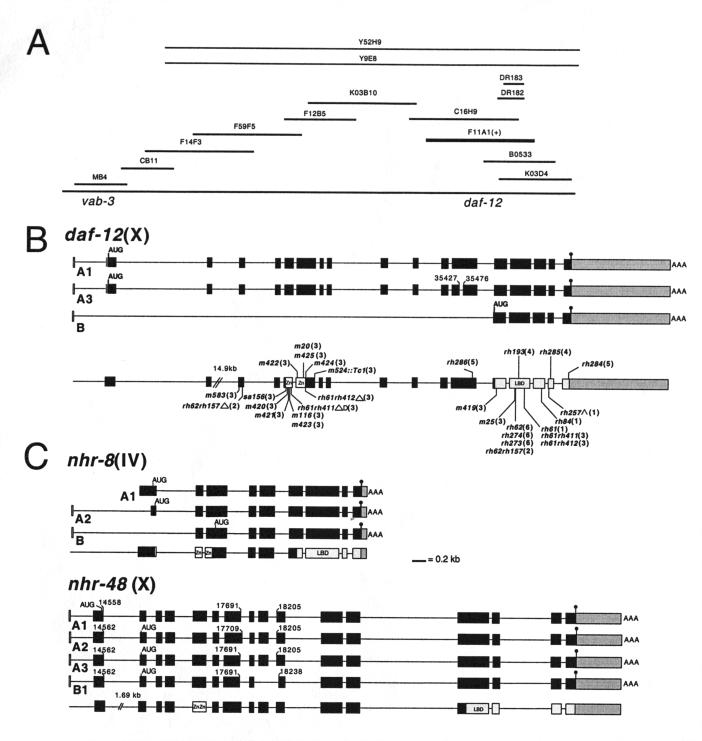
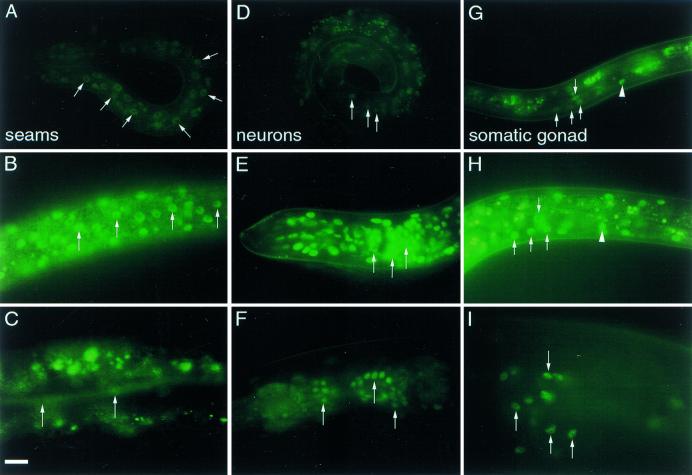
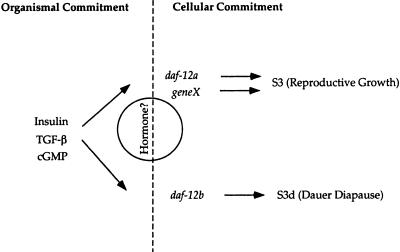
The daf-12 gene acts at the convergence of pathways regulating larval diapause, developmental age, and adult longevity in Caenorhabditis elegans. It encodes a nuclear receptor most closely related to two C. elegans receptors, NHR-8 and NHR-48, Drosophila DHR96, and vertebrate vitamin D and pregnane-X receptors. daf-12 has three predicted protein isoforms, two of which contain DNA- and ligand-binding domains, and one of which contains the ligand-binding domain only. Mutations cluster in DNA- and ligand-binding domains, but correspond to distinct phenotypic classes. DAF-12 is expressed widely in target tissues from embryo to adult, but is upregulated during midlarval stages. In the adult, expression persists in nervous system and somatic gonad, two tissues that regulate adult longevity. We propose that DAF-12 integrates hormonal signals in cellular targets to coordinate major life history traits.
Figures



References
-
- Ambros V. A hierarchy of regulatory genes controls a larva-to-adult developmental switch in C. elegans. Cell. 1989;57:49–57. - PubMed
-
- Ambros V. Heterochronic genes. In: Riddle DL, Blumenthal T, Meyer B, Priess J, editors. C. elegans II. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1997. pp. 501–518. - PubMed
-
- Ambros V, Horvitz HR. Heterochronic mutants of the nematode Caenorhabditis elegans. Science. 1984;226:409–416. - PubMed
-
- Antebi A, Culotti JG, Hedgecock EM. daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development. 1998;125:1191–1205. - PubMed
-
- Antebi A, Norris C, Hedgecock E, Garriga G. Cell and growth cone migrations. In: Riddle DL, Blumenthal T, Meyer B, Priess J, editors. C. elegans II. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1997. pp. 583–609. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous