

Functional significance of the interaction of hepatitis A virus RNA with glyceraldehyde 3-phosphate dehydrogenase (GAPDH): opposing effects of GAPDH and polypyrimidine tract binding protein on internal ribosome entry site function
- PMID: 10864658
- PMCID: PMC112154
- DOI: 10.1128/jvi.74.14.6459-6468.2000
Functional significance of the interaction of hepatitis A virus RNA with glyceraldehyde 3-phosphate dehydrogenase (GAPDH): opposing effects of GAPDH and polypyrimidine tract binding protein on internal ribosome entry site function
Abstract
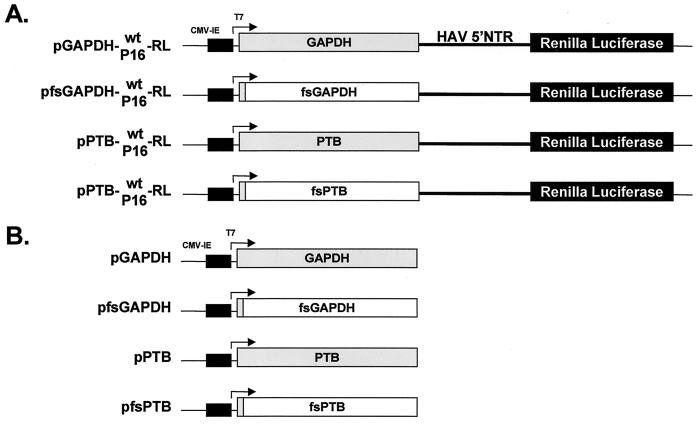
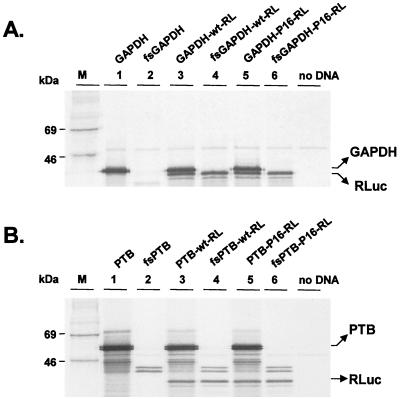
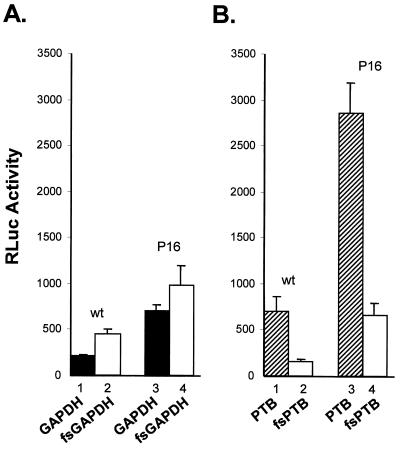
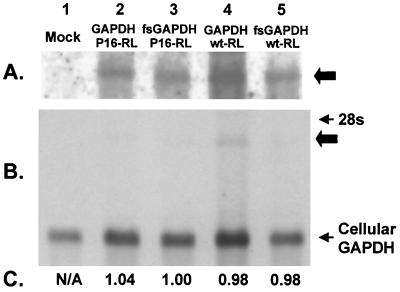
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), a cellular enzyme involved in glycolysis, binds specifically to several viral RNAs, but the functional significance of this interaction is uncertain. Both GAPDH and polypyrimidine tract binding protein (PTB) bind to overlapping sites in stem-loop IIIa of the internal ribosome entry site (IRES) of Hepatitis A virus (HAV), a picornavirus. Since the binding of GAPDH destabilizes the RNA secondary structure, we reasoned that GAPDH may suppress the ability of the IRES to direct cap-independent translation, making its effects antagonistic to the translation-enhancing activity of PTB (D. E. Schultz, C. C. Hardin, and S. M. Lemon, J. Biol. Chem. 271:14134-14142, 1996). To test this hypothesis, we constructed plasmids containing a dicistronic transcriptional unit in which the HAV IRES was placed between an upstream GAPDH-coding sequence and a downstream Renilla luciferase (RLuc) sequence. Transfection with this plasmid results in overexpression of GAPDH and in RLuc production as a measure of IRES activity. RLuc activity was compared with that from a control, null-expression plasmid that was identical except for a frameshift mutation within the 5' GAPDH coding sequence. In transfection experiments, GAPDH overexpression significantly suppressed HAV IRES activity in BSC-1 and FRhK-4 cells but not in Huh-7 cells, which have a significantly greater cytoplasmic abundance of PTB. GAPDH suppression of HAV translation was greater with the wild-type HAV IRES than with the IRES from a cell culture-adapted virus (HM175/P16) that has reproducibly higher basal translational activity in BSC-1 cells. Stem-loop IIIa RNA from the latter IRES had significantly lower affinity for GAPDH in filter binding experiments. Thus, the binding of GAPDH to the IRES of HAV suppresses cap-independent viral translation in vivo in African green monkey kidney cells. The enhanced replication capacity of cell culture-adapted HAV in such cells may be due in part to reduced affinity of the viral IRES for GAPDH.
Figures


References
-
- Borman A, Howell M T, Patton J G, Jackson R J. The involvement of a spliceosome component in internal initiation of human rhinovirus RNA translation. J Gen Virol. 1993;74:1775–1788. - PubMed
-
- Borovjagin A, Pestova T, Shatsky I. Pyrimidine tract binding protein strongly stimulates in vitro encephalomyocarditis virus RNA translation at the level of preinitiation complex formation. FEBS Lett. 1994;351:299–302. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
