

3'-End stem-loops of the subviral RNAs associated with turnip crinkle virus are involved in symptom modulation and coat protein binding
- PMID: 10864666
- PMCID: PMC112162
- DOI: 10.1128/jvi.74.14.6528-6537.2000
3'-End stem-loops of the subviral RNAs associated with turnip crinkle virus are involved in symptom modulation and coat protein binding
Abstract
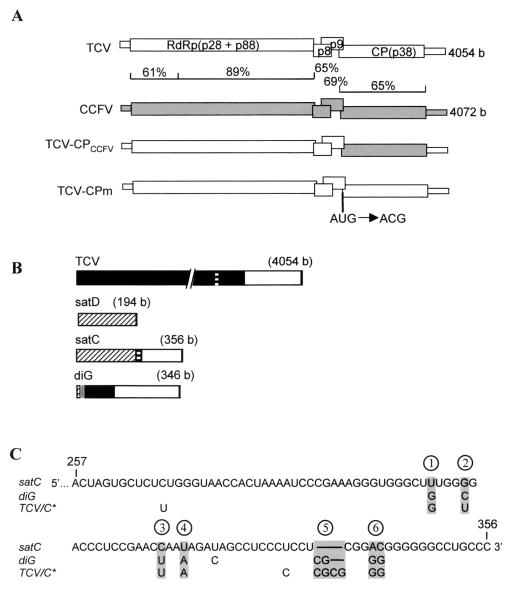
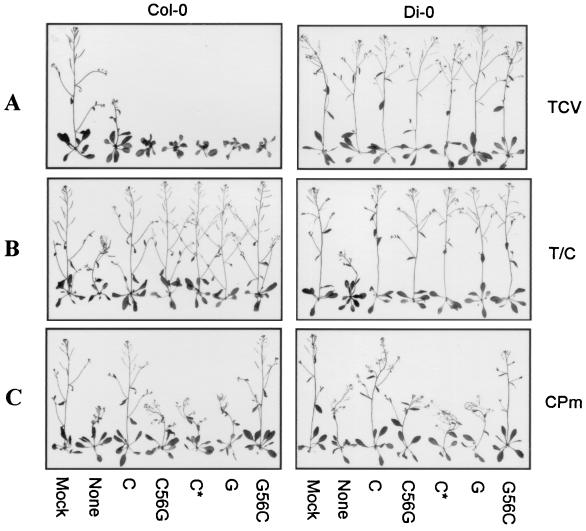
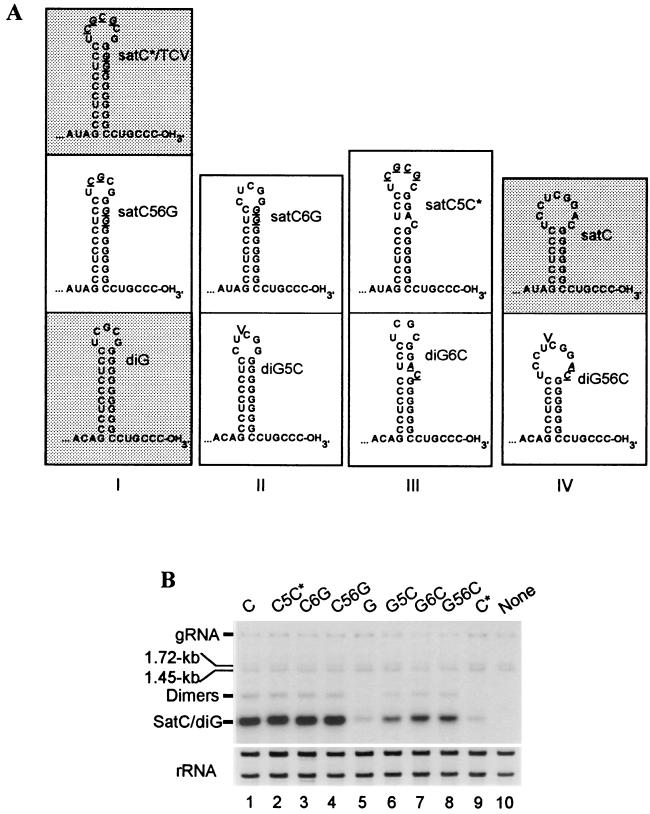
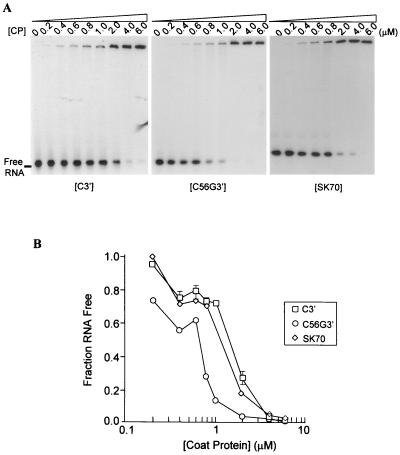
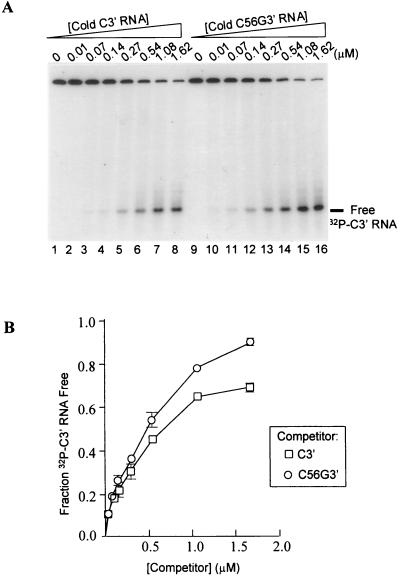
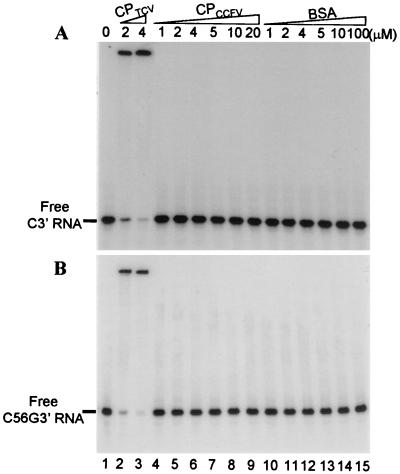
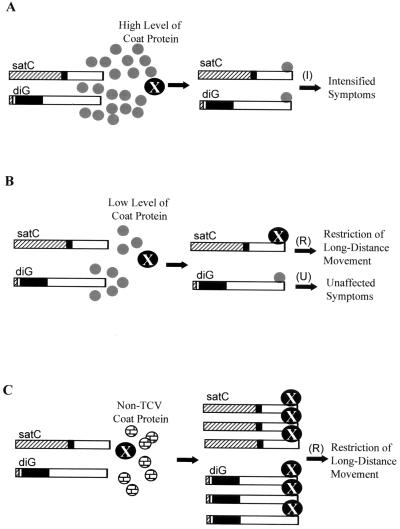
Many plant RNA viruses are associated with one or more subviral RNAs. Two subviral RNAs, satellite RNA C (satC) and defective interfering RNA G (diG) intensify the symptoms of their helper, turnip crinkle virus (TCV). However, when the coat protein (CP) of TCV was replaced with that of the related Cardamine chlorotic fleck virus (CCFV), both subviral RNAs attenuated symptoms of the hybrid virus TCV-CP(CCFV). In contrast, when the translation initiation codon of the TCV CP was altered to ACG and reduced levels of CP were synthesized, satC attenuated symptoms while diG neither intensified nor attenuated symptoms. The determinants for this differential symptom modulation were previously localized to the 3'-terminal 100 bases of the subviral RNAs, which contain six positional differences (Q. Kong, J.-W. Oh, C. D. Carpenter, and A. E. Simon, Virology 238:478-485, 1997). In the current study, we have determined that certain sequences within the 3'-terminal stem-loop structures of satC and diG, which also serve as promoters for complementary strand synthesis, are critical for symptom modulation. Furthermore, the ability to attenuate symptoms was correlated with weakened binding of TCV CP to the hairpin structure.
Figures
Similar articles
-
The coat protein of turnip crinkle virus is involved in subviral RNA-mediated symptom modulation and accumulation.Virology. 1997 Nov 24;238(2):478-85. doi: 10.1006/viro.1997.8853. Virology. 1997. PMID: 9400620
-
Analysis of cis-acting sequences involved in plus-strand synthesis of a turnip crinkle virus-associated satellite RNA identifies a new carmovirus replication element.Virology. 2000 Mar 15;268(2):345-54. doi: 10.1006/viro.1999.0153. Virology. 2000. PMID: 10704342
-
Symptom attenuation by a satellite RNA in vivo is dependent on reduced levels of virus coat protein.Virology. 1999 Jun 20;259(1):234-45. doi: 10.1006/viro.1999.9781. Virology. 1999. PMID: 10364508
-
[Subviral molecules of RNA associated with plant ss(+)RNA viruses].Postepy Biochem. 2006;52(2):212-21. Postepy Biochem. 2006. PMID: 17078511 Review. Polish.
-
Plant virus satellite and defective interfering RNAs: new paradigms for a new century.Annu Rev Phytopathol. 2004;42:415-37. doi: 10.1146/annurev.phyto.42.040803.140402. Annu Rev Phytopathol. 2004. PMID: 15283672 Review.
Cited by
-
Cis and trans requirements for rolling circle replication of a satellite RNA.J Virol. 2004 Mar;78(6):3072-82. doi: 10.1128/jvi.78.6.3072-3082.2004. J Virol. 2004. PMID: 14990726 Free PMC article.
-
Evolution of virus-derived sequences for high-level replication of a subviral RNA.Virology. 2006 Aug 1;351(2):476-88. doi: 10.1016/j.virol.2006.03.011. Epub 2006 May 6. Virology. 2006. PMID: 16682064 Free PMC article.
-
Secondary structural elements within the 3' untranslated region of mouse hepatitis virus strain JHM genomic RNA.J Virol. 2001 Dec;75(24):12105-13. doi: 10.1128/JVI.75.24.12105-12113.2001. J Virol. 2001. PMID: 11711601 Free PMC article.
-
Evolution of a helper virus-derived, ribosome binding translational enhancer in an untranslated satellite RNA of Turnip crinkle virus.Virology. 2011 Oct 10;419(1):10-6. doi: 10.1016/j.virol.2011.07.019. Virology. 2011. PMID: 21862095 Free PMC article.
-
Satellite RNAs and Satellite Viruses of Plants.Viruses. 2009 Dec;1(3):1325-50. doi: 10.3390/v1031325. Epub 2009 Dec 18. Viruses. 2009. PMID: 21994595 Free PMC article.
References
-
- Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Carrington J C, Morris T J, Stockley P G, Harrison S C. Structure and assembly of turnip crinkle virus. IV. Analysis of the coat protein gene and implications of the subunit primary structure. J Mol Biol. 1987;194:265–276. - PubMed
-
- Ferreiro C, Ostrowka K, Lopez-Moya J J, Diaz-Ruiz J R. Nucleotide sequence and symptom modulating analysis of a peanut stunt virus-associated satellite RNA from Poland: high level of sequence identities with the American PSV satellites. Eur J Plant Pathol. 1996;102:779–786.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
