

Analysis of the transmembrane domain of influenza virus neuraminidase, a type II transmembrane glycoprotein, for apical sorting and raft association
- PMID: 10864667
- PMCID: PMC112163
- DOI: 10.1128/jvi.74.14.6538-6545.2000
Analysis of the transmembrane domain of influenza virus neuraminidase, a type II transmembrane glycoprotein, for apical sorting and raft association
Abstract
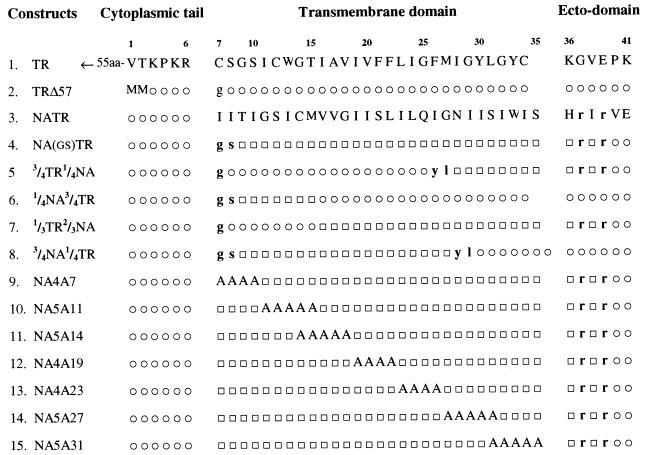
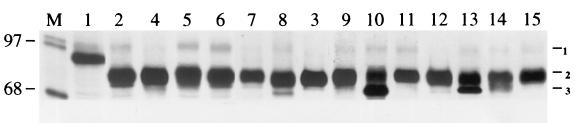
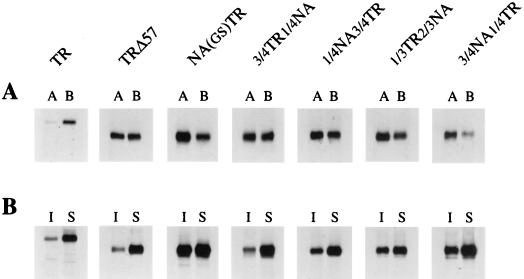
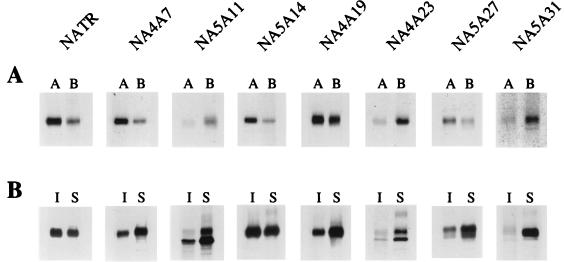
Influenza virus neuraminidase (NA), a type II transmembrane protein, is directly transported to the apical plasma membrane in polarized MDCK cells. Previously, it was shown that the transmembrane domain (TMD) of NA provides a determinant(s) for apical sorting and raft association (A. Kundu, R. T. Avalos, C. M. Sanderson, and D. P. Nayak, J. Virol. 70:6508-6515, 1996). In this report, we have analyzed the sequences in the NA TMD involved in apical transport and raft association by making chimeric TMDs from NA and human transferring receptor (TR) TMDs and by mutating the NA TMD sequences. Our results show that the COOH-terminal half of the NA TMD (amino acids [aa] 19 to 35) was significantly involved in raft association, as determined by Triton X-100 (TX-100) resistance. However, in addition, the highly conserved residues at the extreme NH(2) terminus of the NA TMD were also critical for TX-100 resistance. On the other hand, 19 residues (aa 9 to 27) at the NH(2) terminus of the NA TMD were sufficient for apical sorting. Amino acid residues 14 to 18 and 27 to 31 had the least effect on apical transport, whereas mutations in the amino acid residues 11 to 13, 23 to 26, and 32 to 35 resulted in altered polarity for the mutant proteins. These results indicated that multiple regions in the NA TMD were involved in apical transport. Furthermore, these results support the idea that the signals for apical sorting and raft association, although residing in the NA TMD, are not identical and vary independently and that the NA TMD also possesses an apical determinant(s) which can interact with apical sorting machineries outside the lipid raft.
Figures

References
-
- Brown D A, Rose J K. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
