

Alternative splicing in the cytoplasmic II-III loop of the N-type Ca channel alpha 1B subunit: functional differences are beta subunit-specific
- PMID: 10864934
- PMCID: PMC6772276
- DOI: 10.1523/JNEUROSCI.20-13-04769.2000
Alternative splicing in the cytoplasmic II-III loop of the N-type Ca channel alpha 1B subunit: functional differences are beta subunit-specific
Abstract
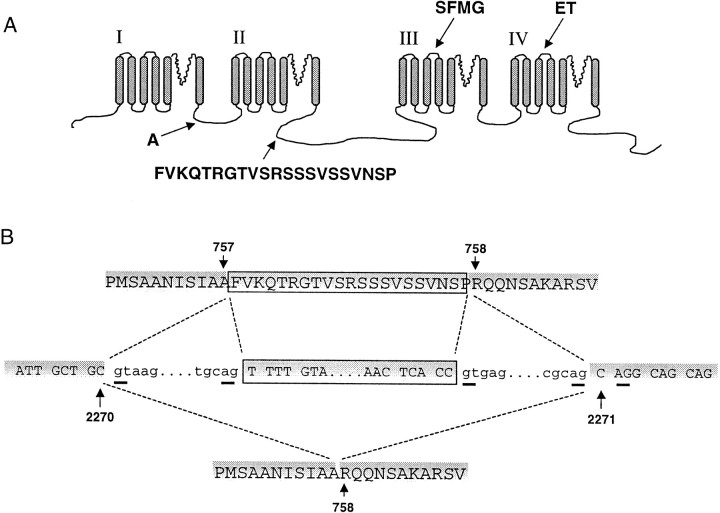
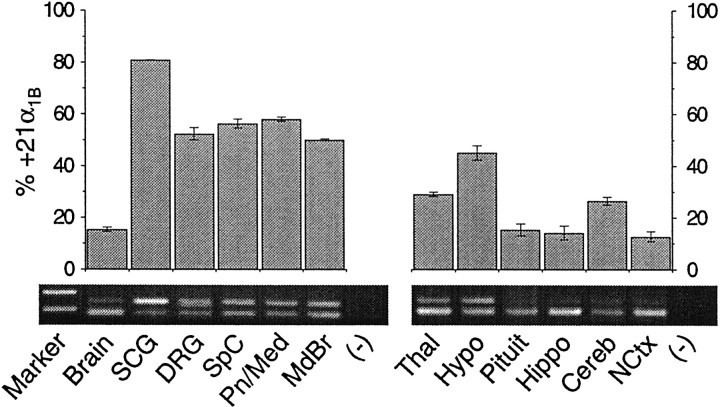
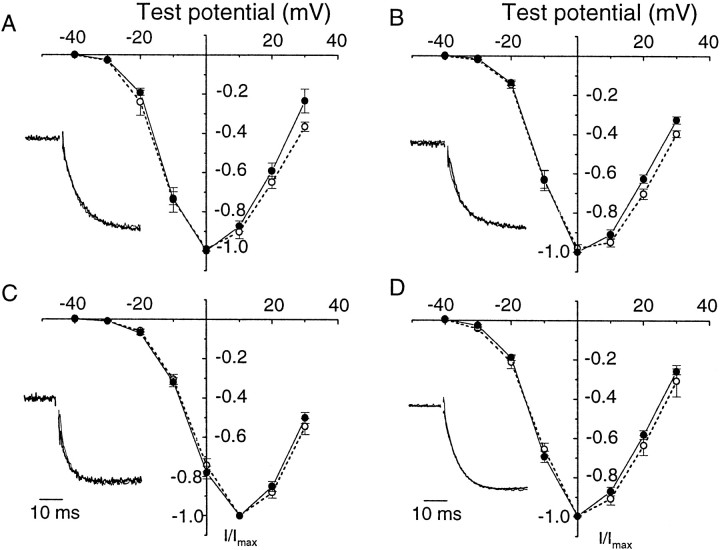
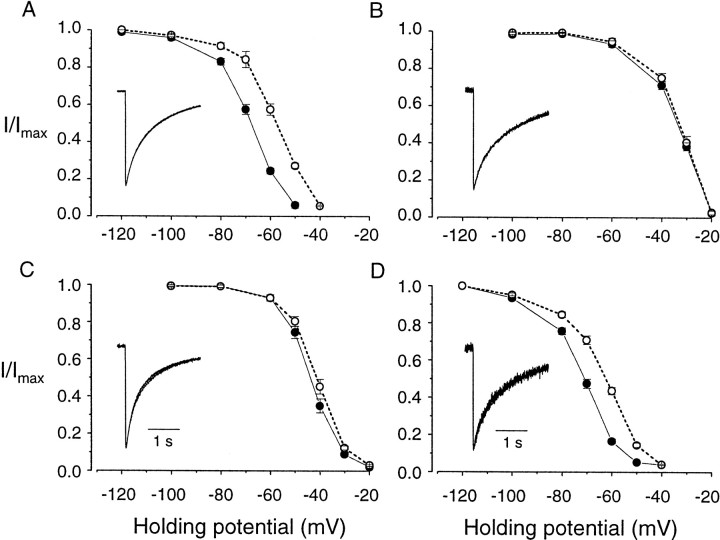
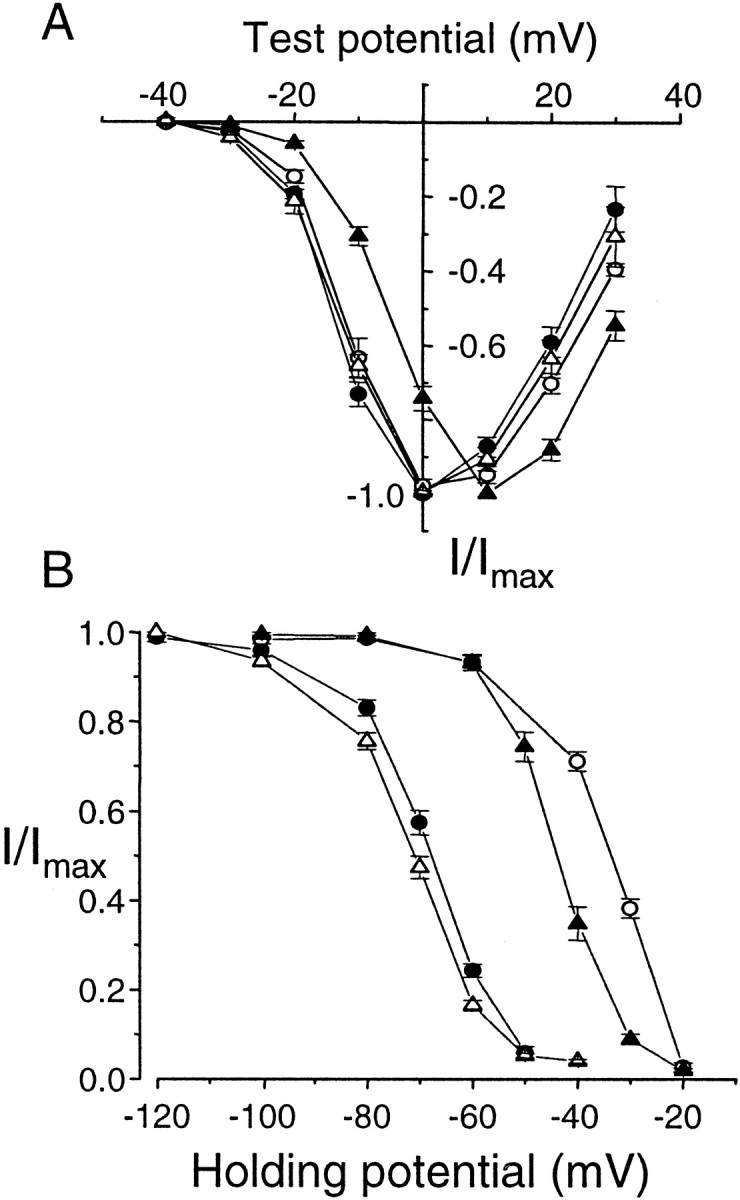
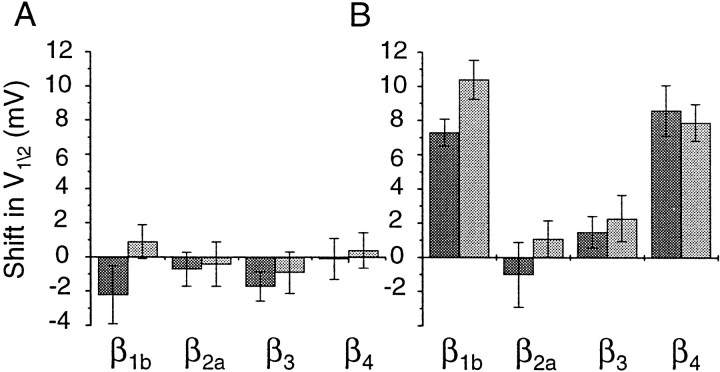
Structural diversity of voltage-gated Ca channels underlies much of the functional diversity in Ca signaling in neurons. Alternative splicing is an important mechanism for generating structural variants within a single gene family. In this paper, we show the expression pattern of an alternatively spliced 21 amino acid encoding exon in the II-III cytoplasmic loop region of the N-type Ca channel alpha(1B) subunit and assess its functional impact. Exon-containing alpha(1B) mRNA dominated in sympathetic ganglia and was present in approximately 50% of alpha(1B) mRNA in spinal cord and caudal regions of the brain and in the minority of alpha(1B) mRNA in neocortex, hippocampus, and cerebellum (<20%). The II-III loop exon affected voltage-dependent inactivation of the N-type Ca channel. Steady-state inactivation curves were shifted to more depolarized potentials without affects on either the rate or voltage dependence of channel opening. Differences in voltage-dependent inactivation between alpha(1B) splice variants were most clearly manifested in the presence of Ca channel beta(1b) or beta(4), rather than beta(2a) or beta(3), subunits. Our results suggest that exon-lacking alpha(1B) splice variants that associate with beta(1b) and beta(4) subunits will be susceptible to voltage-dependent inactivation at voltages in the range of neuronal resting membrane potentials (-60 to -80 mV). In contrast, alpha(1B) splice variants that associate with either beta(2a) or beta(3) subunits will be relatively resistant to inactivation at these voltages. The potential to mix and match multiple alpha(1B) splice variants and beta subunits probably represents a mechanism for controlling the plasticity of excitation-secretion coupling at different synapses.
Figures
References
-
- Bean BP. Neurotransmitter inhibition of neuronal calcium currents by changes in channel voltage dependence. Nature. 1989;340:153–156. - PubMed
-
- Bezprozvanny I, Scheller RH, Tsien RW. Functional impact of syntaxin on gating of N-type and Q-type calcium channels. Nature. 1995;378:623–626. - PubMed
-
- Bourinet E, Soong TW, Sutton K, Slaymaker S, Mathews E, Monteil A, Zamponi GW, Nargeot J, Snutch TP. Splicing of alpha 1A subunit gene generates phenotypic variants of P- and Q-type calcium channels. Nat Neurosci. 1999;2:407–415. - PubMed
-
- Castellano A, Wei X, Birnbaumer L, Perez-Reys E. Cloning and expression of a third calcium channel beta subunit. J Biol Chem. 1993a;268:3450–3455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials