

Active zones on motor nerve terminals contain alpha 3beta 1 integrin
- PMID: 10864949
- PMCID: PMC6772282
- DOI: 10.1523/JNEUROSCI.20-13-04912.2000
Active zones on motor nerve terminals contain alpha 3beta 1 integrin
Abstract
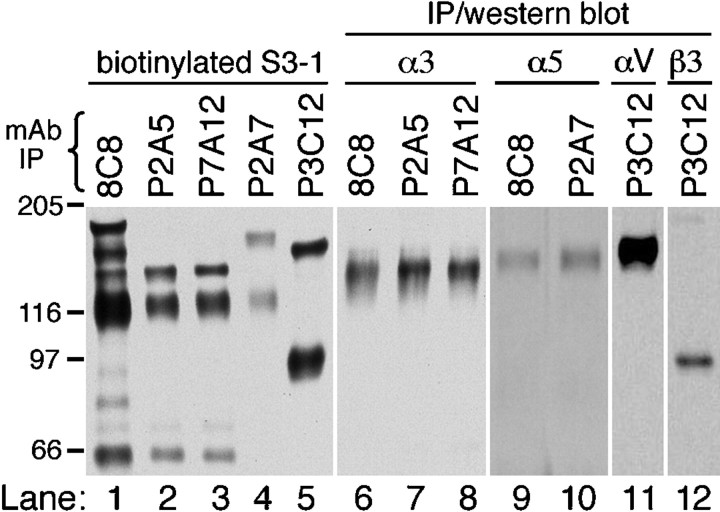
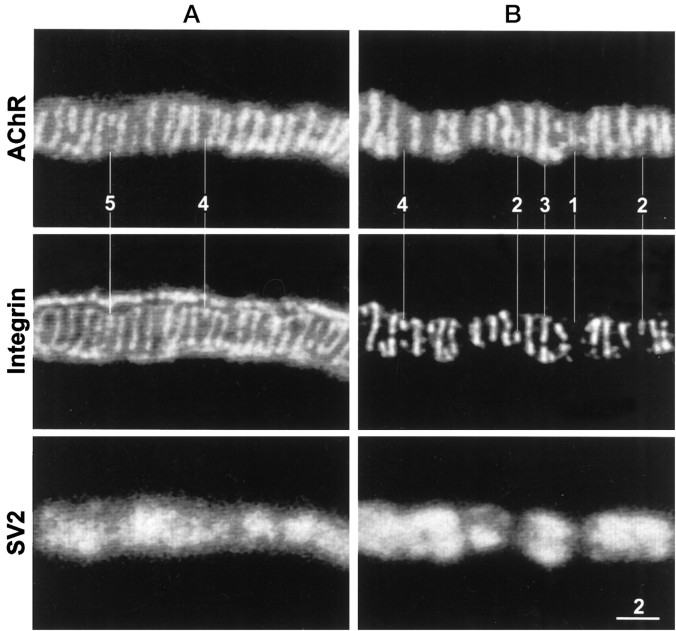
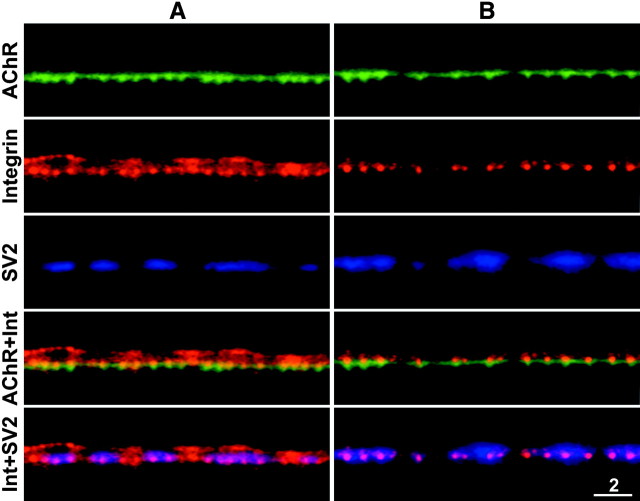
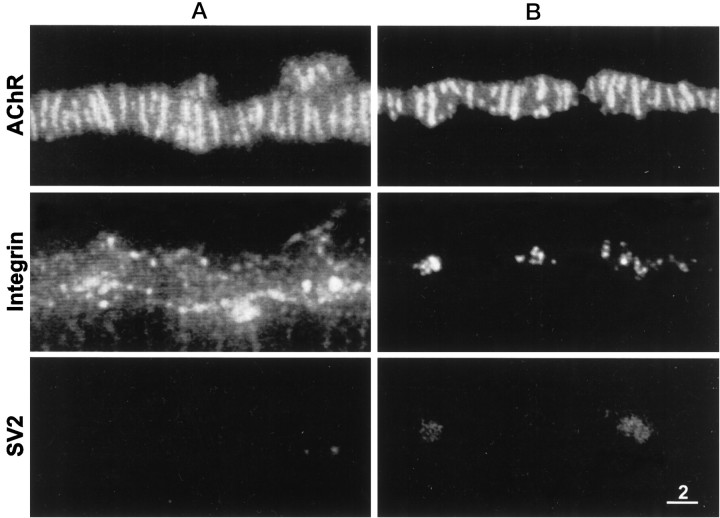
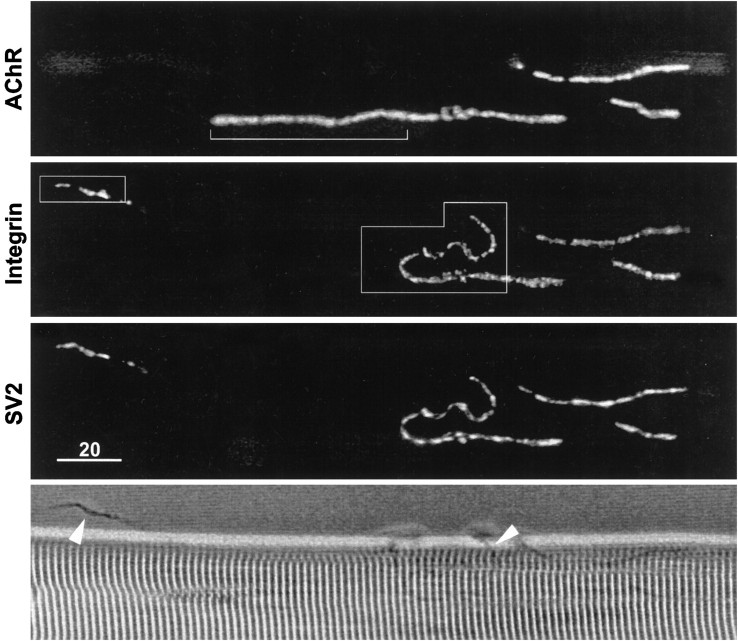
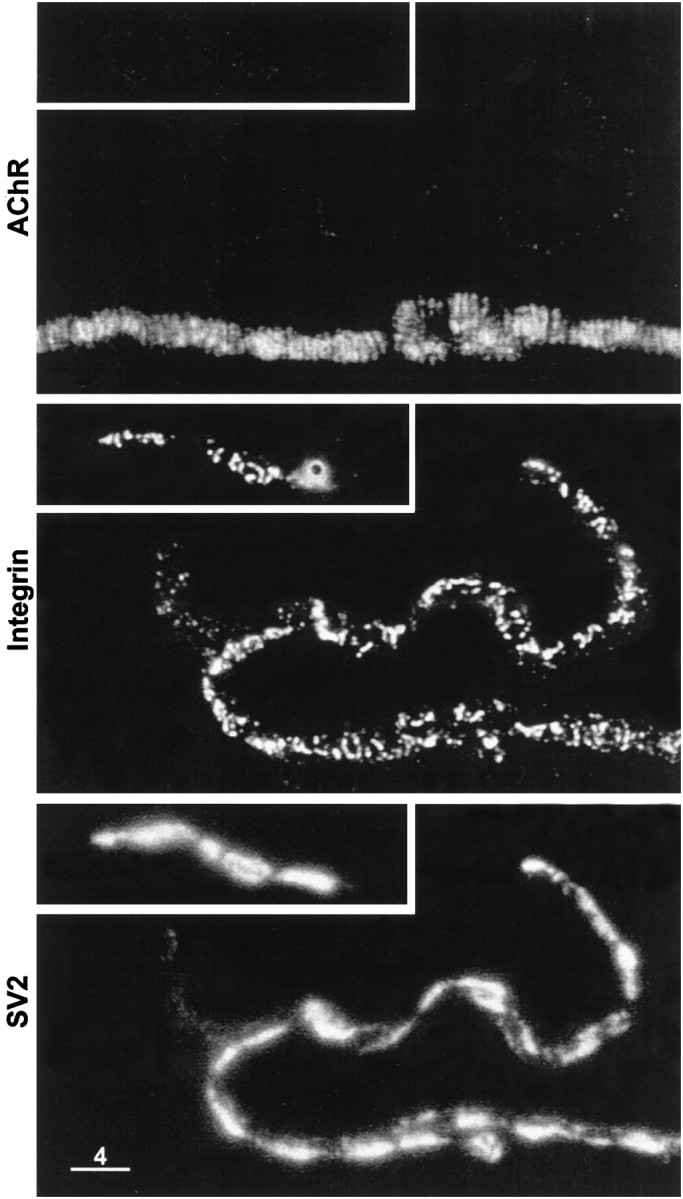
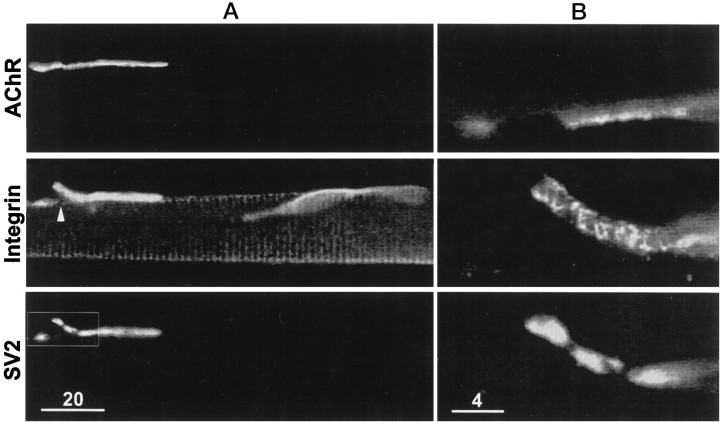
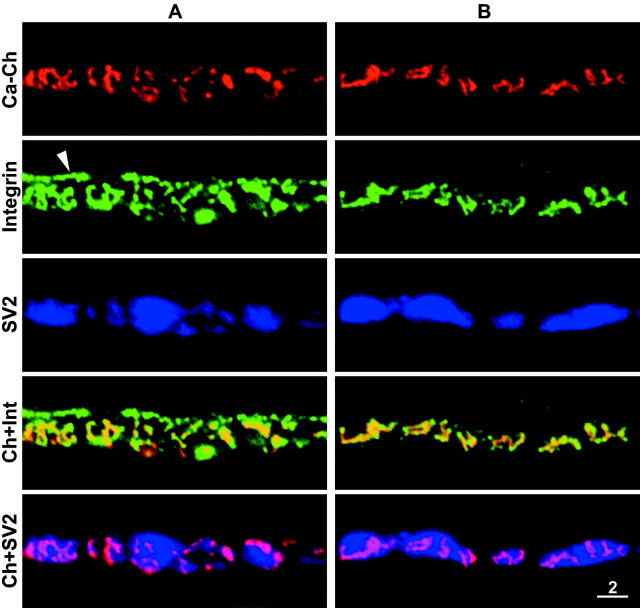
Active zones are the sites along nerve terminals where synaptic vesicles dock and undergo calcium-dependent exocytosis during synaptic transmission. Here we show, by immunofluorescent staining with antibodies generated against Xenopus laevis integrins, that alpha3beta1 integrin is concentrated at the active zones of Xenopus motor nerve terminals. Because integrins can link extracellular matrix molecules to cytoskeletal elements and participate in the formation of signaling complexes, the localization of integrin at active zones suggests that it may play a role in the adhesion of the nerve terminals to the synaptic basal lamina, in the formation and maintenance of active zones, and in some of the events associated with calcium-dependent exocytosis of neurotransmitter. Our findings also indicate that the integrin composition of the terminal Schwann cells differs from that of the motor nerve terminals, and this may account at least in part for differences in their adhesiveness to the synaptic basal lamina.
Figures
References
-
- Aplin AE, Howe A, Alahari SK, Juliano RL. Signal transduction and signal modulation by cell adhesion receptors: the role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins [Review]. Pharmacol Rev. 1998;50:197–263. - PubMed
-
- Astrow SH, Qiang H, Ko CP. Perisynaptic Schwann cells at neuromuscular junctions revealed by a novel monoclonal antibody. J Neurocytol. 1998;27:667–681. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials