

Shared and unique roles of CAP23 and GAP43 in actin regulation, neurite outgrowth, and anatomical plasticity
- PMID: 10871284
- PMCID: PMC2175140
- DOI: 10.1083/jcb.149.7.1443
Shared and unique roles of CAP23 and GAP43 in actin regulation, neurite outgrowth, and anatomical plasticity
Abstract
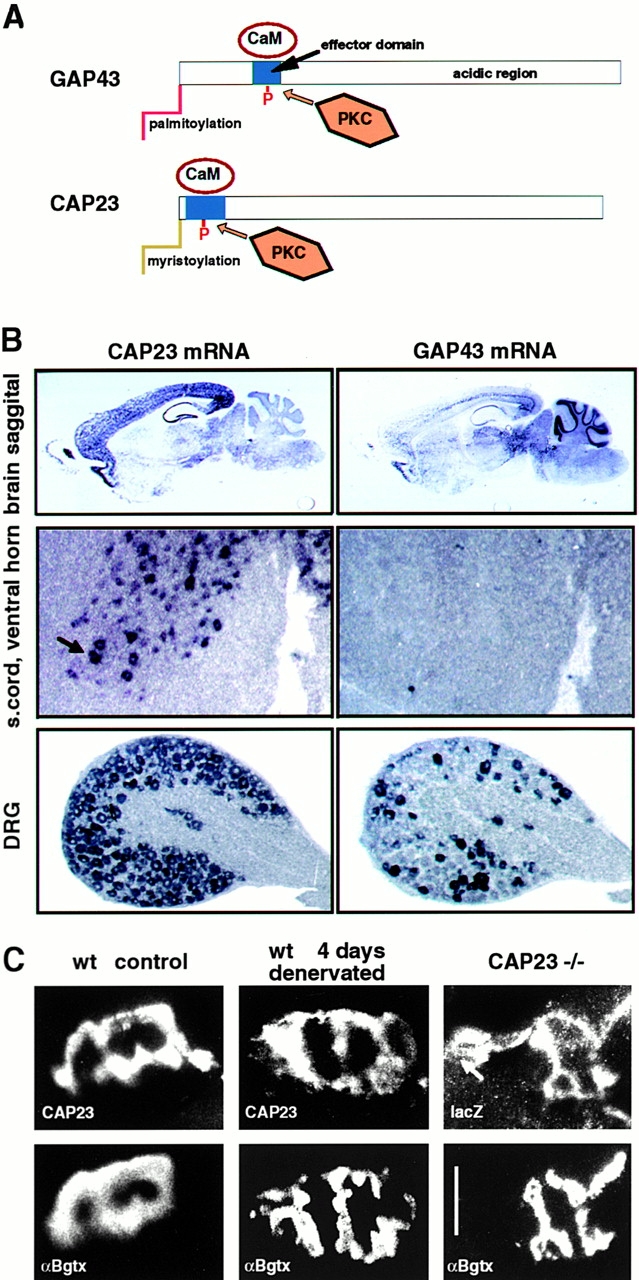
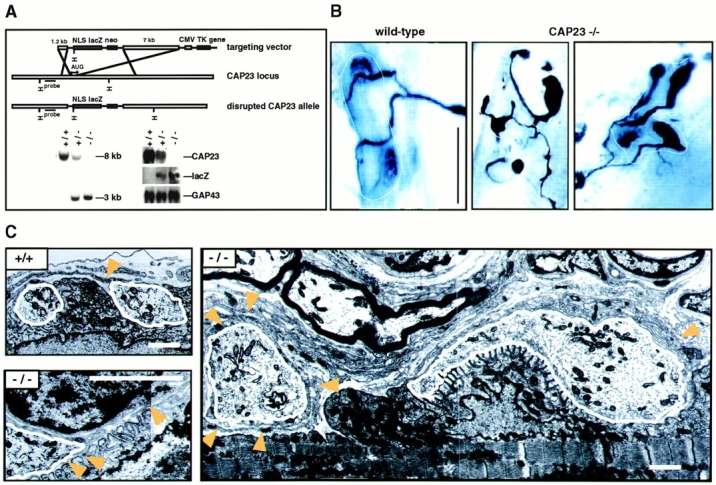
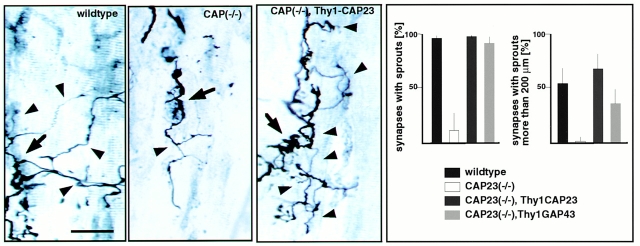
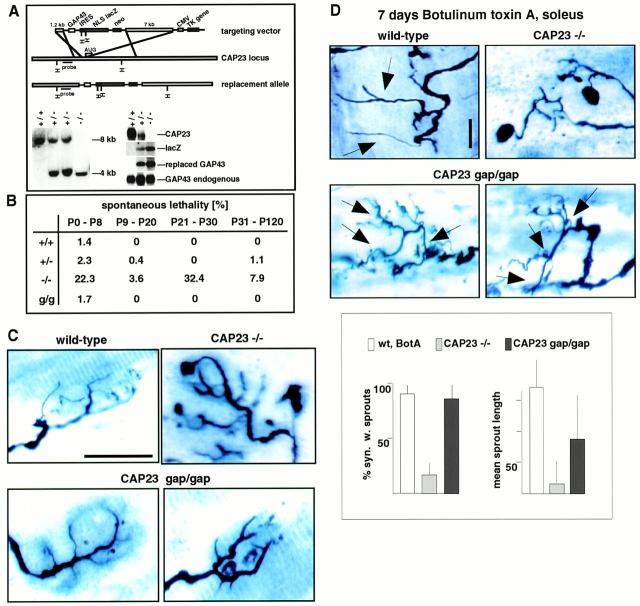
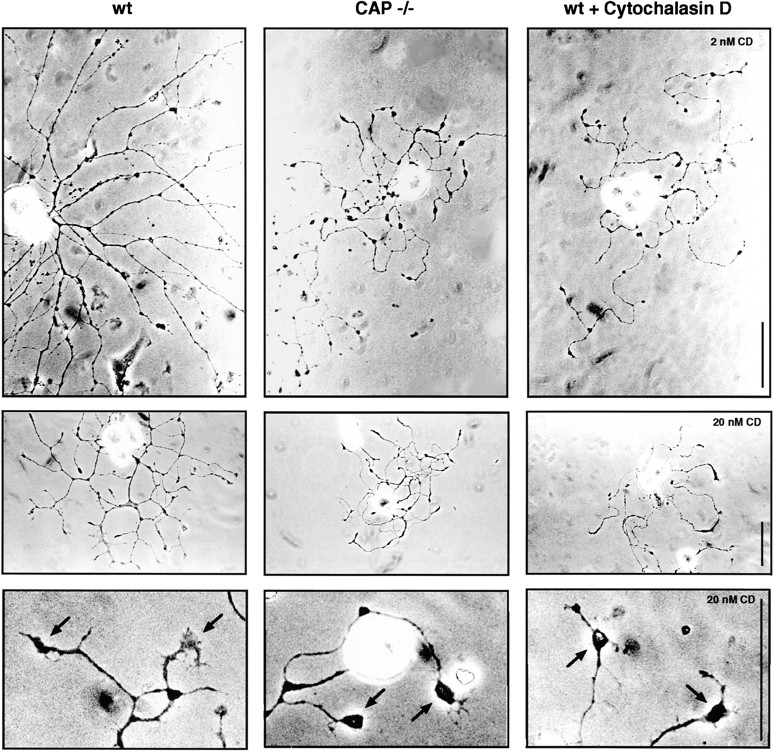
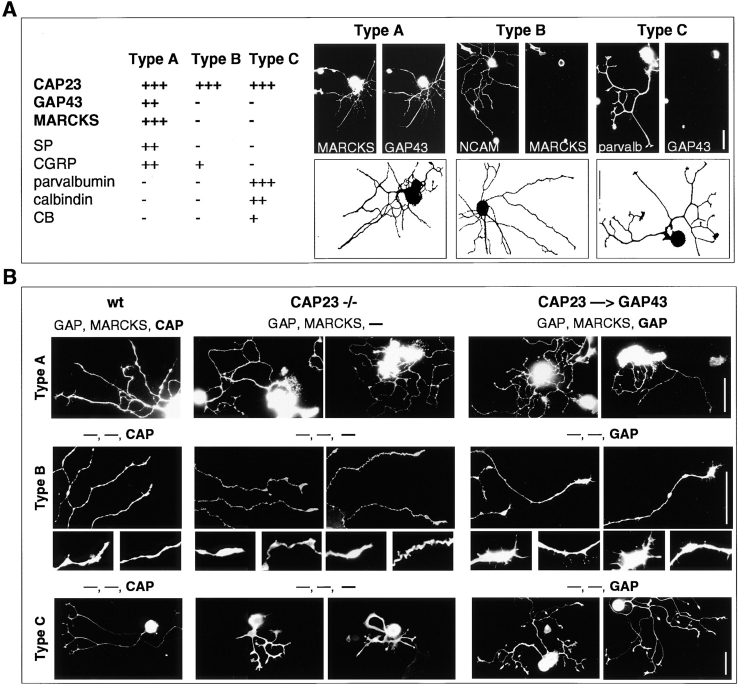
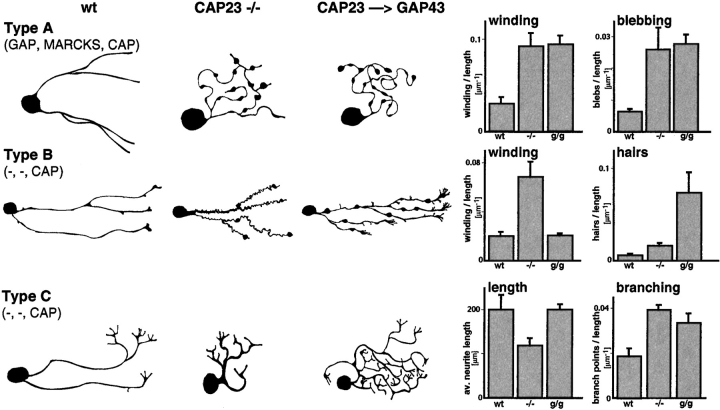
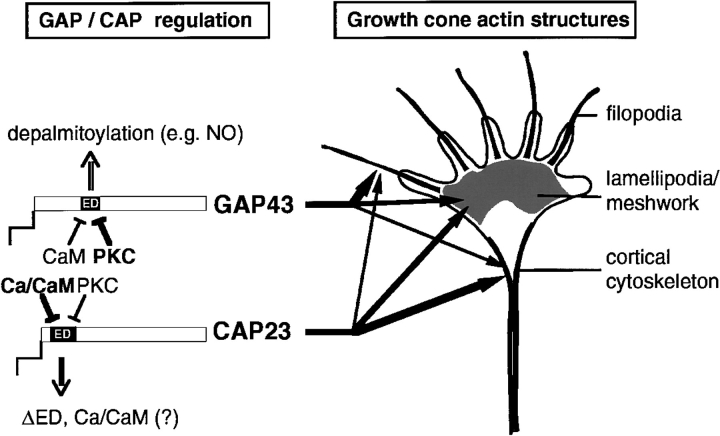
CAP23 is a major cortical cytoskeleton-associated and calmodulin binding protein that is widely and abundantly expressed during development, maintained in selected brain structures in the adult, and reinduced during nerve regeneration. Overexpression of CAP23 in adult neurons of transgenic mice promotes nerve sprouting, but the role of this protein in process outgrowth was not clear. Here, we show that CAP23 is functionally related to GAP43, and plays a critical role to regulate nerve sprouting and the actin cytoskeleton. Knockout mice lacking CAP23 exhibited a pronounced and complex phenotype, including a defect to produce stimulus-induced nerve sprouting at the adult neuromuscular junction. This sprouting deficit was rescued by transgenic overexpression of either CAP23 or GAP43 in adult motoneurons. Knockin mice expressing GAP43 instead of CAP23 were essentially normal, indicating that, although these proteins do not share homologous sequences, GAP43 can functionally substitute for CAP23 in vivo. Cultured sensory neurons lacking CAP23 exhibited striking alterations in neurite outgrowth that were phenocopied by low doses of cytochalasin D. A detailed analysis of such cultures revealed common and unique functions of CAP23 and GAP43 on the actin cytoskeleton and neurite outgrowth. The results provide compelling experimental evidence for the notion that CAP23 and GAP43 are functionally related intrinsic determinants of anatomical plasticity, and suggest that these proteins function by locally promoting subplasmalemmal actin cytoskeleton accumulation.
Figures
References
-
- Aderem A. The MARCKS family of protein kinase-C substrates. Biochem. Soc. Trans. 1995;23:587–591. - PubMed
-
- Aigner, L., S. Arber, J.P. Kapfhammer, T. 0, C. Schneider, F. Botteri, H.-R. Brenner, and P. Caroni. 1995. Overexpression of the neural growth-associated protein GAP-43 induces nerve sprouting in the adult nervous system of transgenic mice. Cell. 83:269–278. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
