

Characterisation of the catalytically active form of RecG helicase
- PMID: 10871364
- PMCID: PMC102718
- DOI: 10.1093/nar/28.12.2324
Characterisation of the catalytically active form of RecG helicase
Abstract
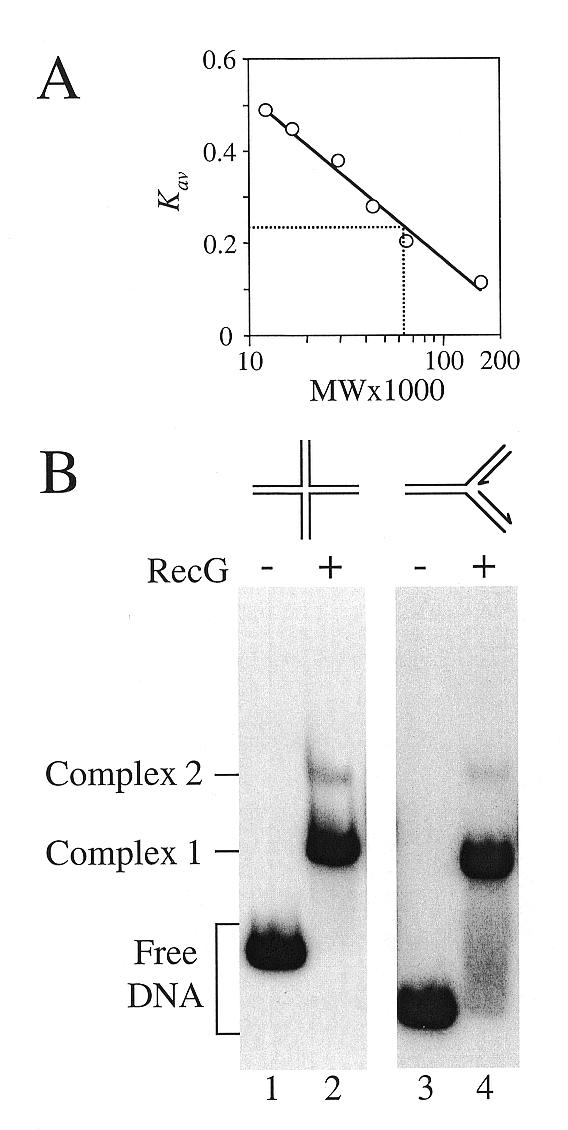
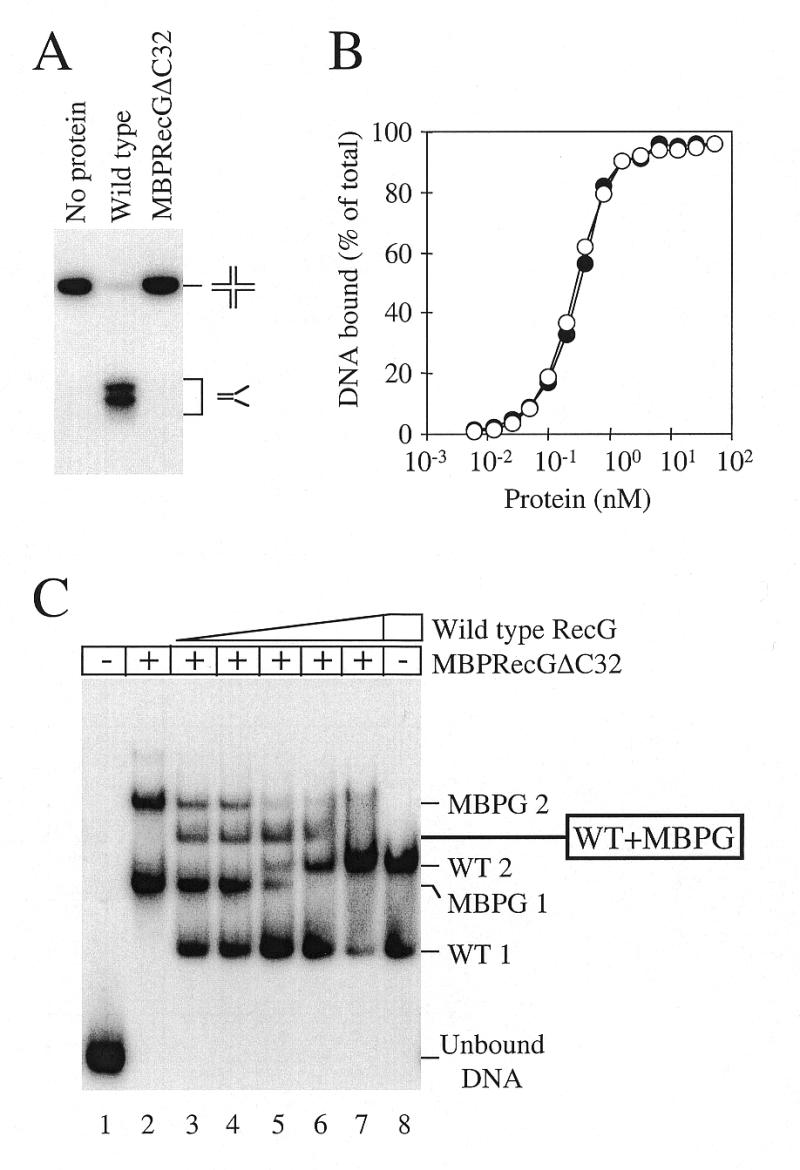
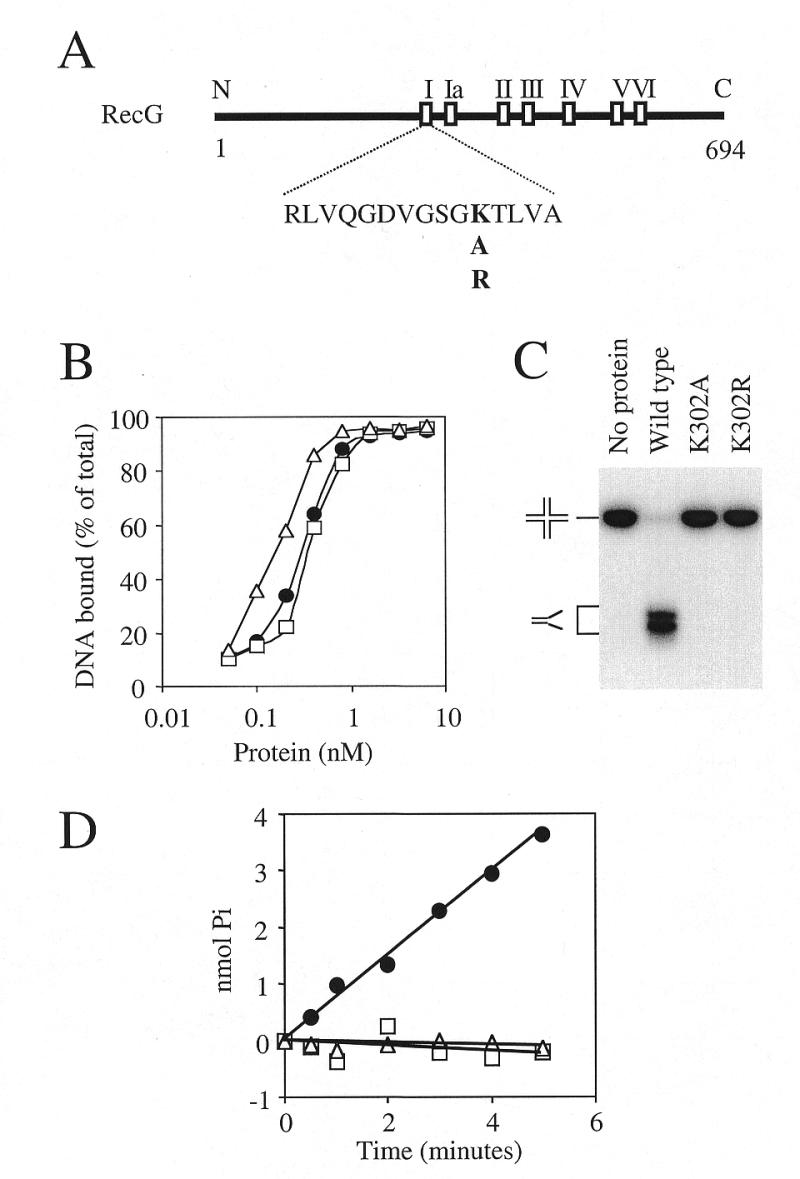
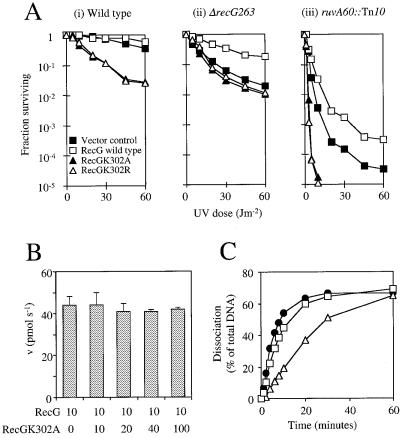
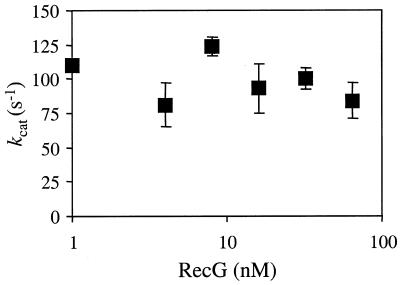
Replication of DNA is fraught with difficulty and chromosomes contain many lesions which may block movement of the replicative machinery. However, several mechanisms to overcome such problems are beginning to emerge from studies with Escherichia coli. An important enzyme in one or more of these mechanisms is the RecG helicase, which may target stalled replication forks to generate a four-stranded (Holliday) junction, thus facilitating repair and/or bypass of the original lesion. To begin to understand how RecG might catalyse regression of fork structures, we have analysed what the catalytically active form of the enzyme may be. We have found that RecG exists as a monomer in solution as measured by gel filtration but when bound to junction DNA the enzyme forms two distinct protein-DNA complexes that contain one and two protein molecules. However, mutant inhibition studies failed to provide any evidence that RecG acts as a multimer in vitro. Additionally, there was no evidence for cooperativity in the junction DNA-stimulated hydrolysis of ATP. These data suggest that RecG functions as a monomer to unwind junction DNA, which supports an 'inchworm' rather than an 'active rolling' mechanism of DNA unwinding. The observed in vivo inhibition of wild-type RecG by mutant forms of the enzyme was attributed to occlusion of the DNA target and correlates with the very low abundance of replication forks within an E.COLI: cell, even during rapid growth.
Figures
References
-
- McGlynn P. and Lloyd,R.G. (2000) Cell, 101, 35–45. - PubMed
-
- Friedberg E.C., Walker,G.C. and Siede,W. (1995) In Friedberg,E.C., Walker,G.C. and Siede,W. (eds), DNA Repair and Mutagenesis. ASM Press, Washington, DC.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
