

Structure-based predictions of Rad1, Rad9, Hus1 and Rad17 participation in sliding clamp and clamp-loading complexes
- PMID: 10871397
- PMCID: PMC102700
- DOI: 10.1093/nar/28.13.2481
Structure-based predictions of Rad1, Rad9, Hus1 and Rad17 participation in sliding clamp and clamp-loading complexes
Abstract
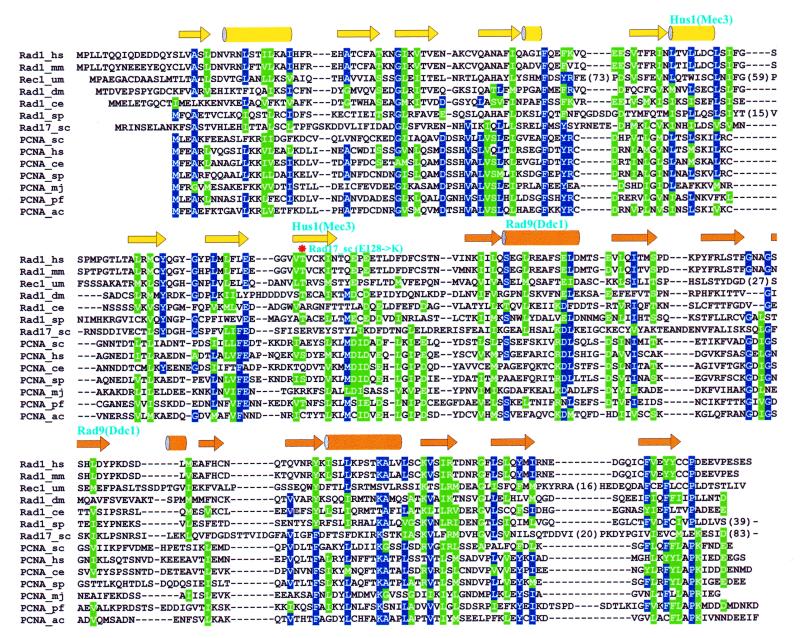
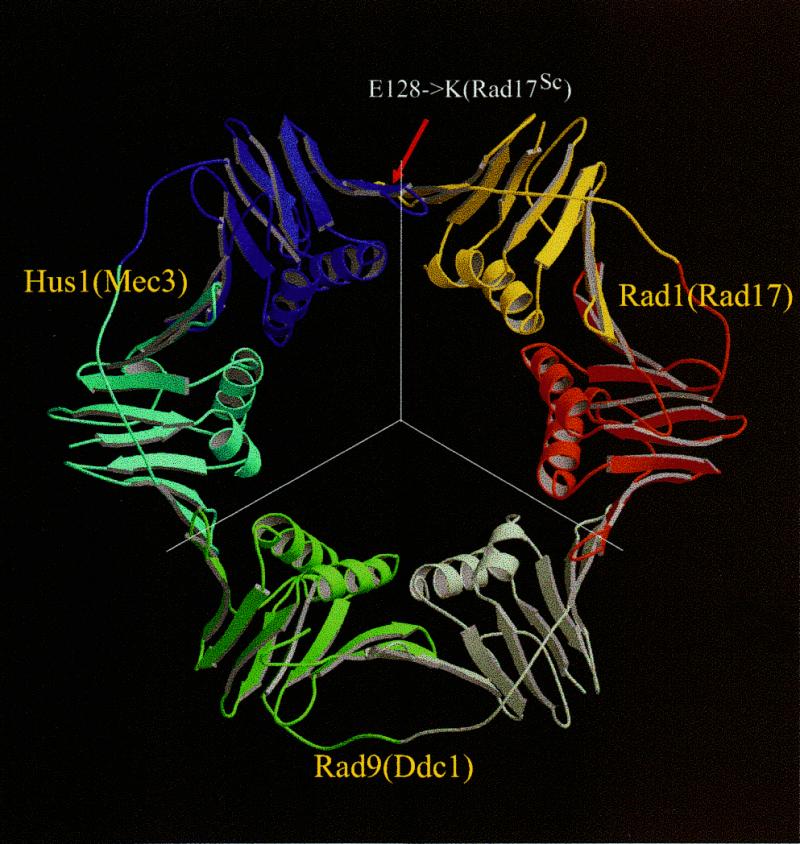
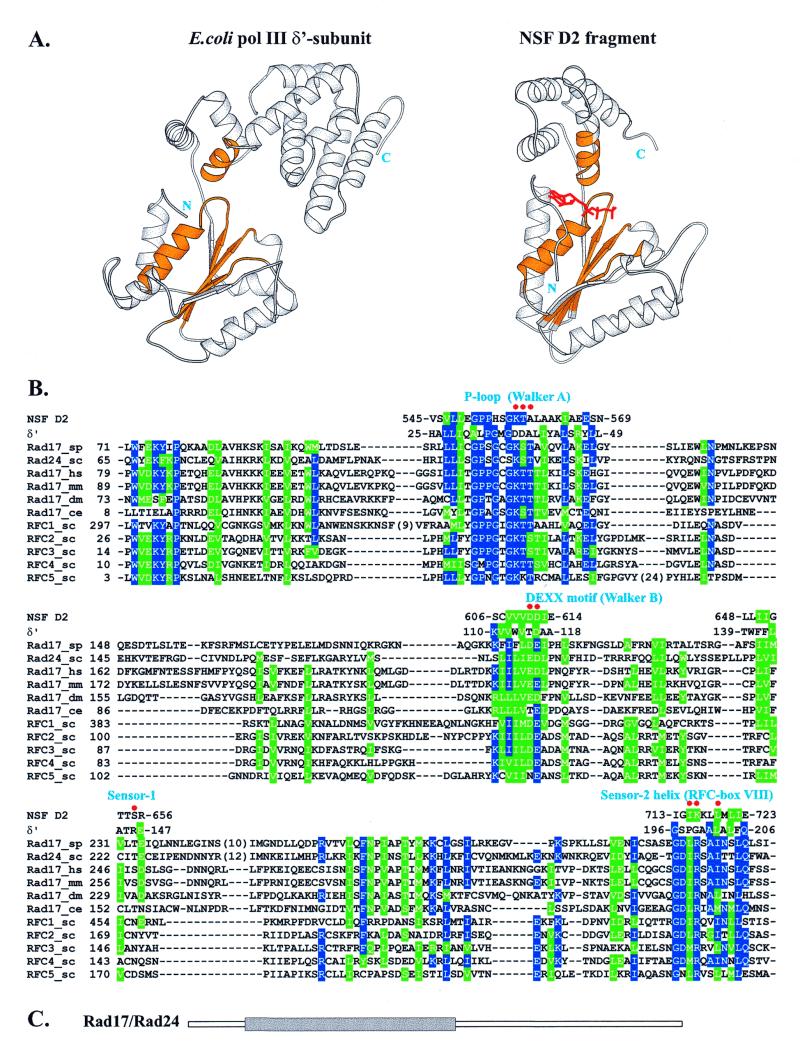
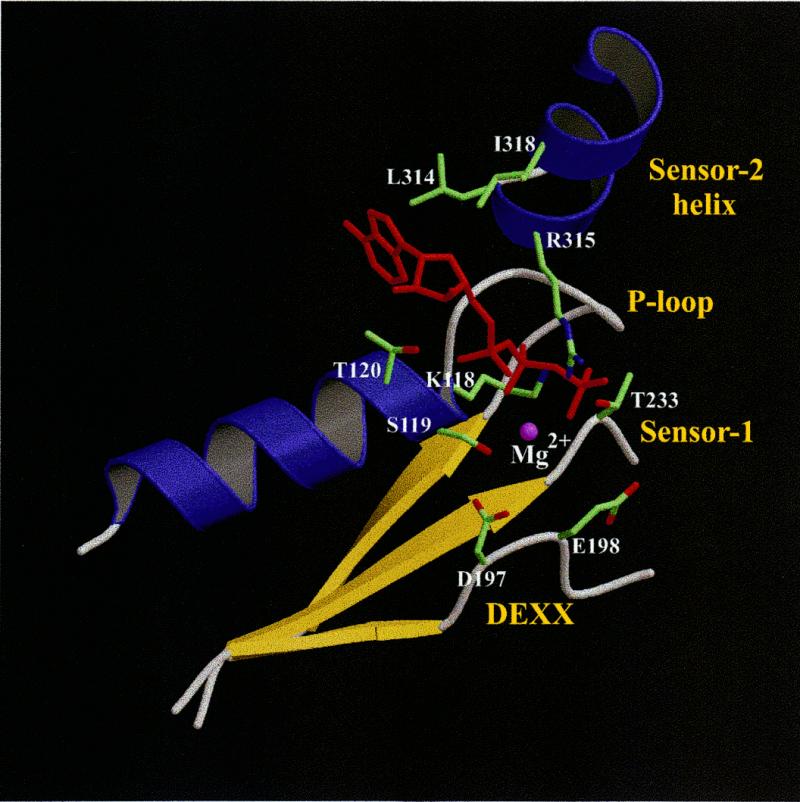
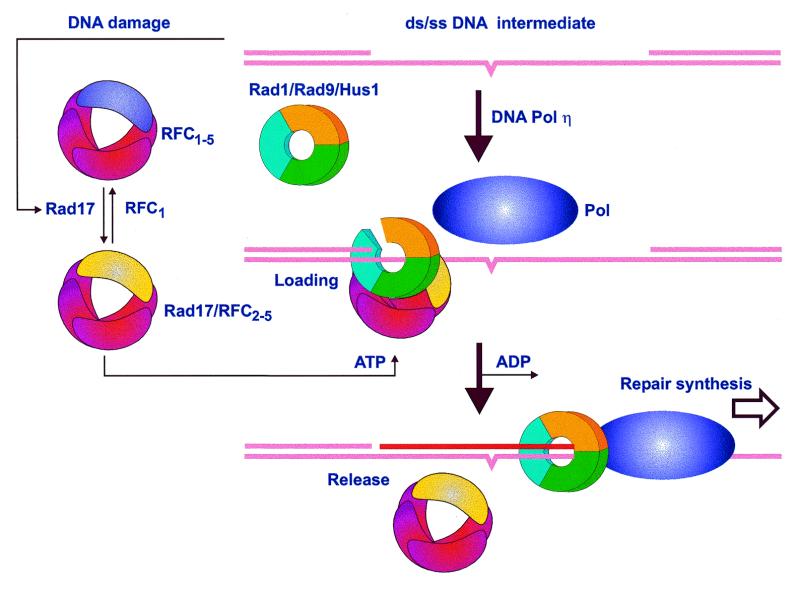
The repair of damaged DNA is coupled to the completion of DNA replication by several cell cycle checkpoint proteins, including, for example, in fission yeast Rad1(Sp), Hus1(Sp), Rad9(Sp) and Rad17(Sp). We have found that these four proteins are conserved with protein sequences throughout eukaryotic evolution. Using computational techniques, including fold recognition, comparative modeling and generalized sequence profiles, we have made high confidence structure predictions for the each of the Rad1, Hus1 and Rad9 protein families (Rad17(Sc), Mec3(Sc) and Ddc1(Sc) in budding yeast, respectively). Each of these families was found to share a common protein fold with that of PCNA, the sliding clamp protein that tethers DNA polymerase to its template. We used previously reported genetic and biochemical data for these proteins from yeast and human cells to predict a heterotrimeric PCNA-like ring structure for the functional Rad1/Rad9/Hus1 complex and to determine their exact order within it. In addition, for each individual protein family, contact regions with neighbors within the PCNA-like ring were identified. Based on a molecular model for Rad17(Sp), we concluded that members of this family, similar to the subunits of the RFC clamp-loading complex, are capable of coupling ATP binding with conformational changes required to load a sliding clamp onto DNA. This model substantiates previous findings regarding the behavior of Rad17 family proteins upon DNA damage and within the RFC complex of clamp-loading proteins.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
