

Gene expression profile in prion protein-deficient fibroblasts in culture
- PMID: 10880376
- PMCID: PMC1850192
- DOI: 10.1016/s0002-9440(10)64517-8
Gene expression profile in prion protein-deficient fibroblasts in culture
Abstract
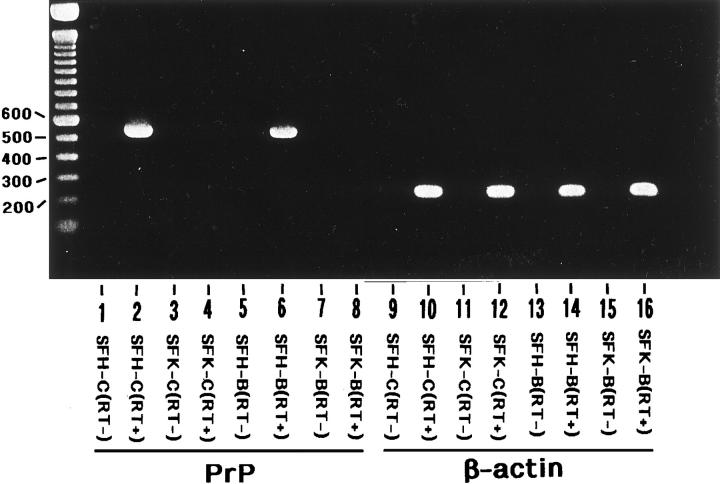
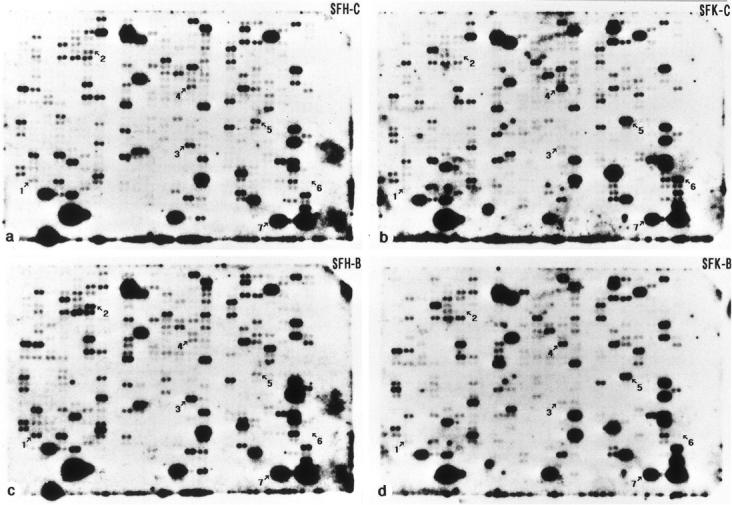
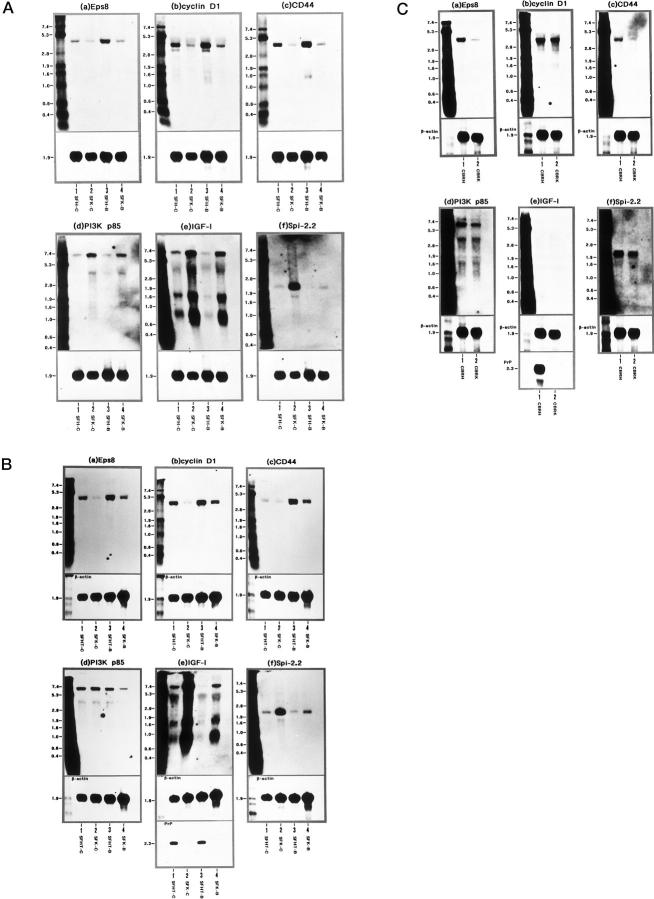
To investigate the physiological function of the cellular isoform of prion protein (PrP(C)), the gene expression profile was studied by analyzing a cDNA expression array containing 597 clones of various functional classes in two distinct skin fibroblast cell lines designated SFK and SFH, established from PrP-deficient (PrP(-)(/-)) mice and PrP(+/+) mice, respectively. The cells were incubated in the culture medium with or without inclusion of basic fibroblast growth factor (bFGF). When SFK cells were compared with SFH cells in untreated conditions, the expression of 15 genes, including those essential for cell proliferation and adhesion, was reduced, whereas the expression of 27 genes, including those involved in the insulin-like growth factor-I (IGF-I) signaling pathway, was elevated. Northern blot analysis verified a significant down-regulation of the receptor tyrosine kinase substrate Eps8, cyclin D1, and CD44 mRNAs, and a substantial up-regulation of phosphatidylinositol 3-kinase p85, IGF-I, and serine protease inhibitor-2.2 mRNAs in SFK cells. The patterns of induction or reduction of gene expression after exposure to bFGF showed considerable overlap between both cell types. Furthermore, both Eps8 and CD44 mRNA levels were reduced greatly in the brain tissues of the cerebrum isolated from the PrP(-)(/-) mice. These results indicate that the disruption of the PrP gene resulted in an aberrant regulation of a battery of genes important for cell proliferation, differentiation, and survival, including those located in the Ras and Rac signaling pathways.
Figures
Similar articles
-
Cultured skin fibroblasts isolated from mice devoid of the prion protein gene express major heat shock proteins in response to heat stress.Exp Neurol. 1998 May;151(1):105-15. doi: 10.1006/exnr.1998.6796. Exp Neurol. 1998. PMID: 9582258
-
Modulation of prion protein gene expression by growth factors in cultured mouse astrocytes and PC-12 cells.Brain Res Mol Brain Res. 1994 Mar;22(1-4):268-74. doi: 10.1016/0169-328x(94)90055-8. Brain Res Mol Brain Res. 1994. PMID: 7912403
-
Developmental influence of the cellular prion protein on the gene expression profile in mouse hippocampus.Physiol Genomics. 2011 Jun 28;43(12):711-25. doi: 10.1152/physiolgenomics.00205.2010. Epub 2011 Mar 15. Physiol Genomics. 2011. PMID: 21406608
-
PrP knock-out and PrP transgenic mice in prion research.Br Med Bull. 2003;66:43-60. doi: 10.1093/bmb/66.1.43. Br Med Bull. 2003. PMID: 14522848 Review.
-
Molecular biology and transgenetics of prion diseases.Crit Rev Biochem Mol Biol. 1991;26(5-6):397-438. doi: 10.3109/10409239109086789. Crit Rev Biochem Mol Biol. 1991. PMID: 1684745 Review.
Cited by
-
Loss of prion protein induces a primed state of type I interferon-responsive genes.PLoS One. 2017 Jun 26;12(6):e0179881. doi: 10.1371/journal.pone.0179881. eCollection 2017. PLoS One. 2017. PMID: 28651013 Free PMC article.
-
The Prion Protein Regulates Synaptic Transmission by Controlling the Expression of Proteins Key to Synaptic Vesicle Recycling and Exocytosis.Mol Neurobiol. 2019 May;56(5):3420-3436. doi: 10.1007/s12035-018-1293-4. Epub 2018 Aug 20. Mol Neurobiol. 2019. PMID: 30128651
-
Cellular prion protein and caveolin-1 interaction in a neuronal cell line precedes Fyn/Erk 1/2 signal transduction.J Biomed Biotechnol. 2006;2006(5):69469. doi: 10.1155/JBB/2006/69469. J Biomed Biotechnol. 2006. PMID: 17489019 Free PMC article.
-
Transcriptomic analysis brings new insight into the biological role of the prion protein during mouse embryogenesis.PLoS One. 2011;6(8):e23253. doi: 10.1371/journal.pone.0023253. Epub 2011 Aug 15. PLoS One. 2011. PMID: 21858045 Free PMC article.
-
Role of hypoxia‑mediated cellular prion protein functional change in stem cells and potential application in angiogenesis (Review).Mol Med Rep. 2017 Nov;16(5):5747-5751. doi: 10.3892/mmr.2017.7387. Epub 2017 Aug 29. Mol Med Rep. 2017. PMID: 28901450 Free PMC article.
References
-
- Bendheim PE, Brown HR, Rudelli RD, Scala LJ, Goller NL, Wen GY, Kascsak RJ, Cashman NR, Bolton DC: Nearly ubiquitous tissue distribution of the scrapie agent precursor protein. Neurology 1992, 42:149-156 - PubMed
-
- Brown DR, Qin K, Herms JW, Madlung A, Manson J, Strome R, Fraser PE, Kruck T, von Bohlen A, Schulz-Schaeffer W, Giese A, Westaway D, Kretzschmar H: The cellular prion protein binds copper in vivo. Nature 1997, 390:684-687 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
