

BMP-2 antagonists emerge from alterations in the low-affinity binding epitope for receptor BMPR-II
- PMID: 10880444
- PMCID: PMC313944
- DOI: 10.1093/emboj/19.13.3314
BMP-2 antagonists emerge from alterations in the low-affinity binding epitope for receptor BMPR-II
Abstract
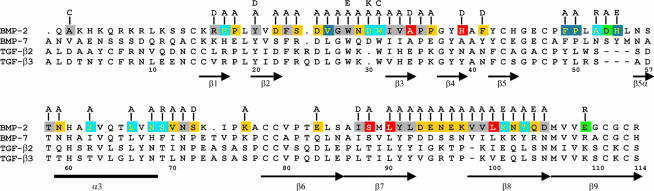
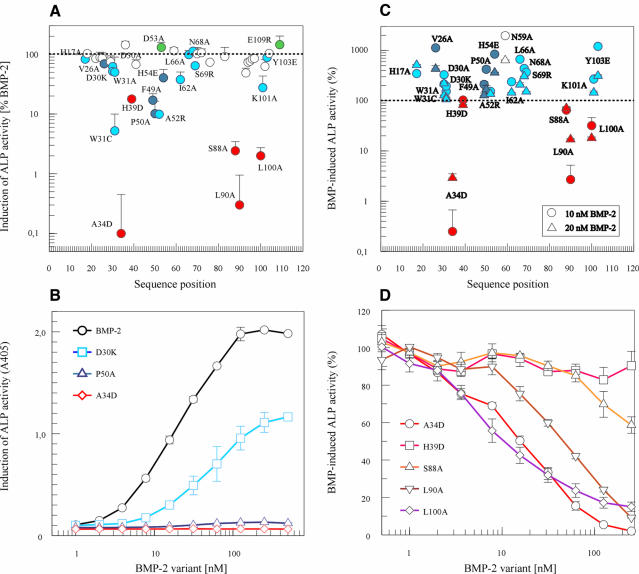
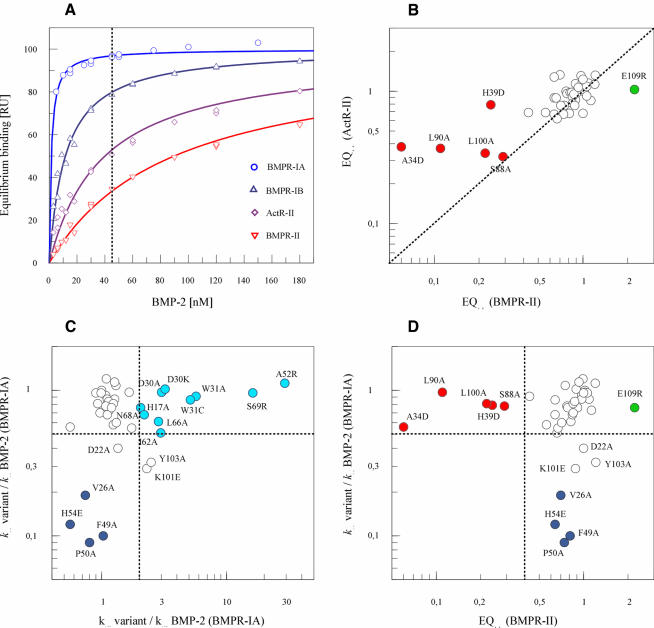
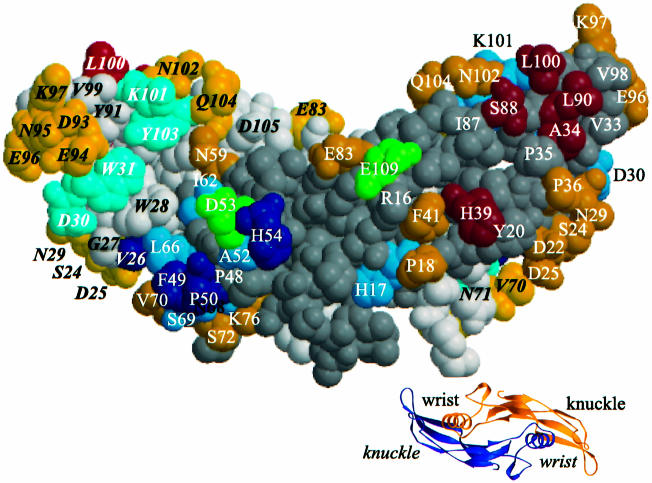
Bone morphogenetic protein-2 (BMP-2) induces bone formation and regeneration in adult vertebrates and regulates important developmental processes in all animals. BMP-2 is a homodimeric cysteine knot protein that, as a member of the transforming growth factor-beta (TGF-beta) superfamily, signals by oligomerizing type I and type II receptor serine-kinases in the cell membrane. The binding epitopes of BMP-2 for BMPR-IA (type I) and BMPR-II or ActR-II (type II) were characterized using BMP-2 mutant proteins for analysis of interactions with receptor ectodomains. A large epitope 1 for high-affinity BMPR-IA binding was detected spanning the interface of the BMP-2 dimer. A smaller epitope 2 for the low-affinity binding of BMPR-II was found to be assembled by determinants of a single monomer. Symmetry-related pairs of the two juxtaposed epitopes occur near the BMP-2 poles. Mutations in both epitopes yielded variants with reduced biological activity in C2C12 cells; however, only epitope 2 variants behaved as antagonists partially or completely inhibiting BMP-2 activity. These findings provide a framework for the molecular description of receptor recognition and activation in the BMP/TGF-beta superfamily.
Figures
References
-
- Black J. (1989) Drugs from emasculated hormones: the principle of syntopic antagonism. Science, 245, 486–493. - PubMed
-
- Blessing M., Schirmacher,P. and Kaiser,S. (1996) Overexpression of bone morphogenetic protein-6 (BMP-6) in the epidermis of transgenic mice: inhibition or stimulation of proliferation depending on the pattern of transgene expression and formation of psoriatic lesions. J. Cell Biol., 135, 227–239. - PMC - PubMed
-
- Chalaux E., Lopez Rovira,T., Rosa,J.L., Bartrons,R. and Ventura,F. (1998) JunB is involved in the inhibition of myogenic differentiation by bone morphogenetic protein-2. J. Biol. Chem., 273, 537–543. - PubMed
-
- Chen Y., Riese,M.J., Killinger,M.A. and Hoffmann,F.M. (1998) A genetic screen for modifiers of Drosophila decapentaplegic signaling identifies mutations in punt, Mothers against dpp and the BMP-7 homologue, 60A. Development, 125, 1759–1768. - PubMed
-
- Cunningham B.C. and Wells,J.A. (1993) Comparison of a structural and a functional epitope [published erratum appears in J. Mol. Biol.,237, 513]. J. Mol. Biol., 234, 554–563. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
