

Arginines 29 and 59 of elongation factor G are important for GTP hydrolysis or translocation on the ribosome
- PMID: 10880458
- PMCID: PMC313957
- DOI: 10.1093/emboj/19.13.3458
Arginines 29 and 59 of elongation factor G are important for GTP hydrolysis or translocation on the ribosome
Abstract
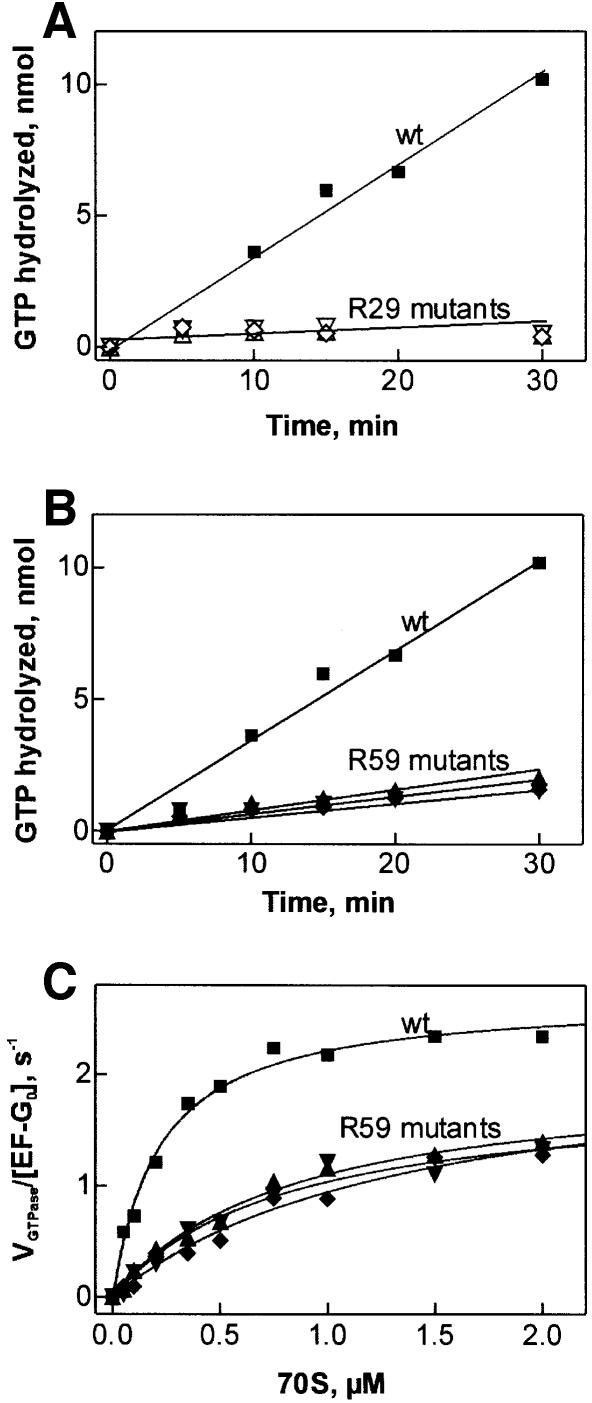
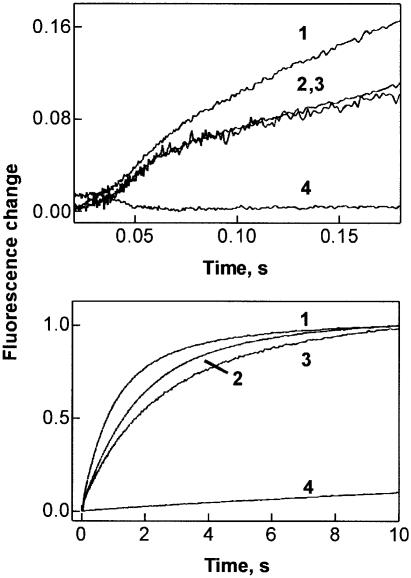
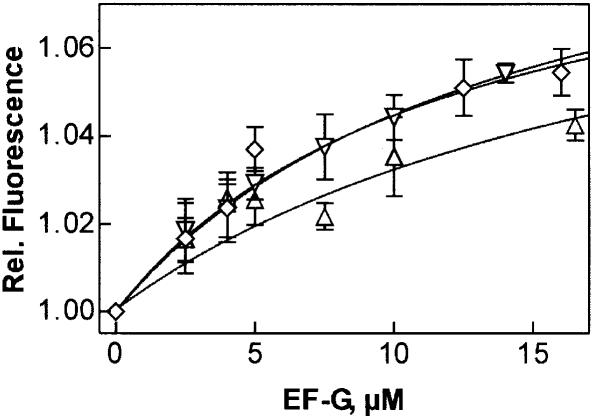
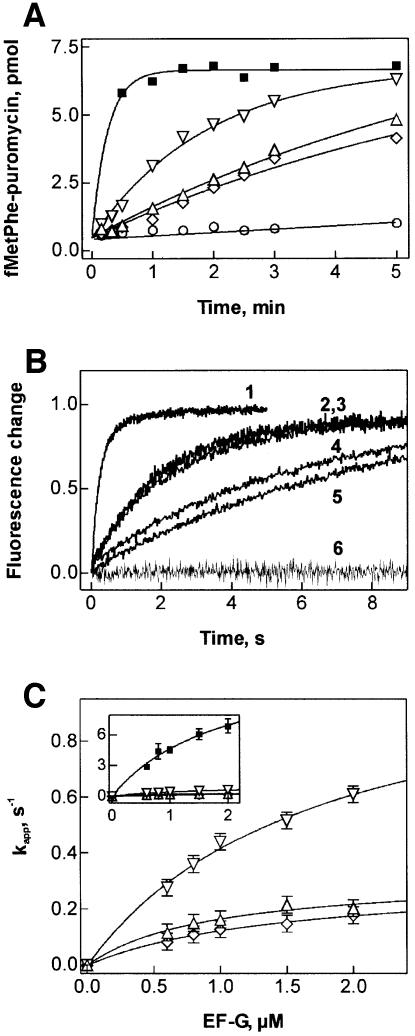
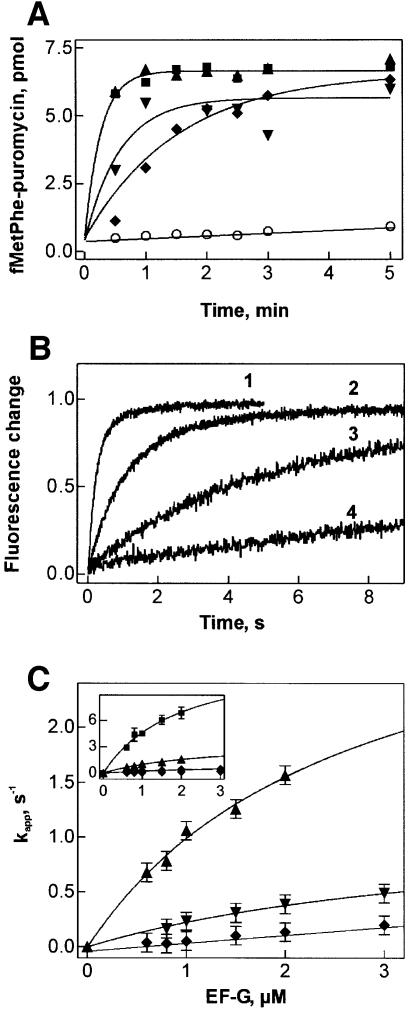
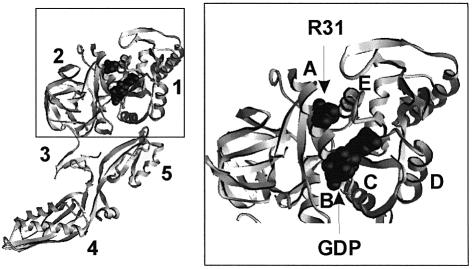
GTP hydrolysis by elongation factor G (EF-G) is essential for the translocation step in protein elongation. The low intrinsic GTPase activity of EF-G is strongly stimulated by the ribosome. Here we show that a conserved arginine, R29, of Escherichia coli EF-G is crucial for GTP hydrolysis on the ribosome, but not for GTP binding or ribosome interaction, suggesting that it may be directly involved in catalysis. Another conserved arginine, R59, which is homologous to the catalytic arginine of G(alpha) proteins, is not essential for GTP hydrolysis, but influences ribosome binding and translocation. These results indicate that EF-G is similar to other GTPases in that an arginine residue is required for GTP hydrolysis, although the structural changes leading to GTPase activation are different.
Figures
Similar articles
-
Role of domains 4 and 5 in elongation factor G functions on the ribosome.J Mol Biol. 2000 Jul 21;300(4):951-61. doi: 10.1006/jmbi.2000.3886. J Mol Biol. 2000. PMID: 10891280
-
Domain III of elongation factor G from Thermus thermophilus is essential for induction of GTP hydrolysis on the ribosome.J Biol Chem. 2000 Nov 17;275(46):35820-4. doi: 10.1074/jbc.M002656200. J Biol Chem. 2000. PMID: 10940297
-
EF-G Activation by Phosphate Analogs.J Mol Biol. 2016 May 22;428(10 Pt B):2248-58. doi: 10.1016/j.jmb.2016.03.032. Epub 2016 Apr 8. J Mol Biol. 2016. PMID: 27063503 Free PMC article.
-
Synchronous tRNA movements during translocation on the ribosome are orchestrated by elongation factor G and GTP hydrolysis.Bioessays. 2014 Oct;36(10):908-18. doi: 10.1002/bies.201400076. Epub 2014 Aug 13. Bioessays. 2014. PMID: 25118068 Review.
-
EF-G and EF4: translocation and back-translocation on the bacterial ribosome.Nat Rev Microbiol. 2014 Feb;12(2):89-100. doi: 10.1038/nrmicro3176. Epub 2013 Dec 23. Nat Rev Microbiol. 2014. PMID: 24362468 Review.
Cited by
-
Characterization of the snowy cotyledon 1 mutant of Arabidopsis thaliana: the impact of chloroplast elongation factor G on chloroplast development and plant vitality.Plant Mol Biol. 2006 Mar;60(4):507-18. doi: 10.1007/s11103-005-4921-0. Plant Mol Biol. 2006. PMID: 16525888
-
Control of ribosomal subunit rotation by elongation factor G.Science. 2013 Jun 28;340(6140):1235970. doi: 10.1126/science.1235970. Science. 2013. PMID: 23812721 Free PMC article.
-
Optimization of a fluorescent-mRNA based real-time assay for precise kinetic measurements of ribosomal translocation.RNA Biol. 2021 Dec;18(12):2363-2375. doi: 10.1080/15476286.2021.1913312. Epub 2021 May 3. RNA Biol. 2021. PMID: 33938388 Free PMC article.
-
Role and timing of GTP binding and hydrolysis during EF-G-dependent tRNA translocation on the ribosome.Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13670-5. doi: 10.1073/pnas.0606099103. Epub 2006 Aug 29. Proc Natl Acad Sci U S A. 2006. PMID: 16940356 Free PMC article.
-
High-resolution structures of a thermophilic eukaryotic 80S ribosome reveal atomistic details of translocation.Nat Commun. 2022 Jan 25;13(1):476. doi: 10.1038/s41467-022-27967-9. Nat Commun. 2022. PMID: 35079002 Free PMC article.
References
-
- Ahmadian M.R., Stege,P., Scheffzek,K. and Wittinghofer,A. (1997) Confirmation of the arginine-finger hypothesis for the GAP-stimulated GTP-hydrolysis reaction of Ras. Nature Struct. Biol., 4, 686–689. - PubMed
-
- Ban N., Nissen,P., Hansen,J., Capel,M., Moore,P.B. and Steitz,T.A. (1999) Placement of protein and RNA structures into a 5 Å-resolution map of the 50S ribosomal subunit. Nature, 400, 841–847. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
