

Use of inhibitors to evaluate coreceptor usage by simian and simian/human immunodeficiency viruses and human immunodeficiency virus type 2 in primary cells
- PMID: 10888629
- PMCID: PMC112207
- DOI: 10.1128/jvi.74.15.6893-6910.2000
Use of inhibitors to evaluate coreceptor usage by simian and simian/human immunodeficiency viruses and human immunodeficiency virus type 2 in primary cells
Abstract
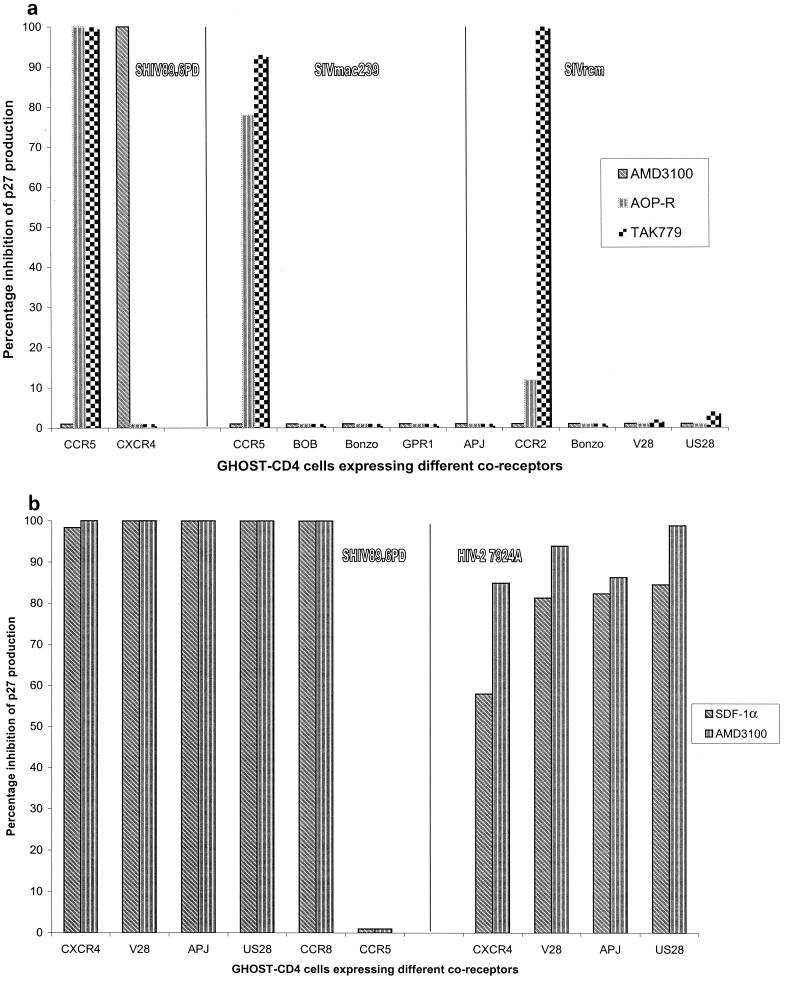
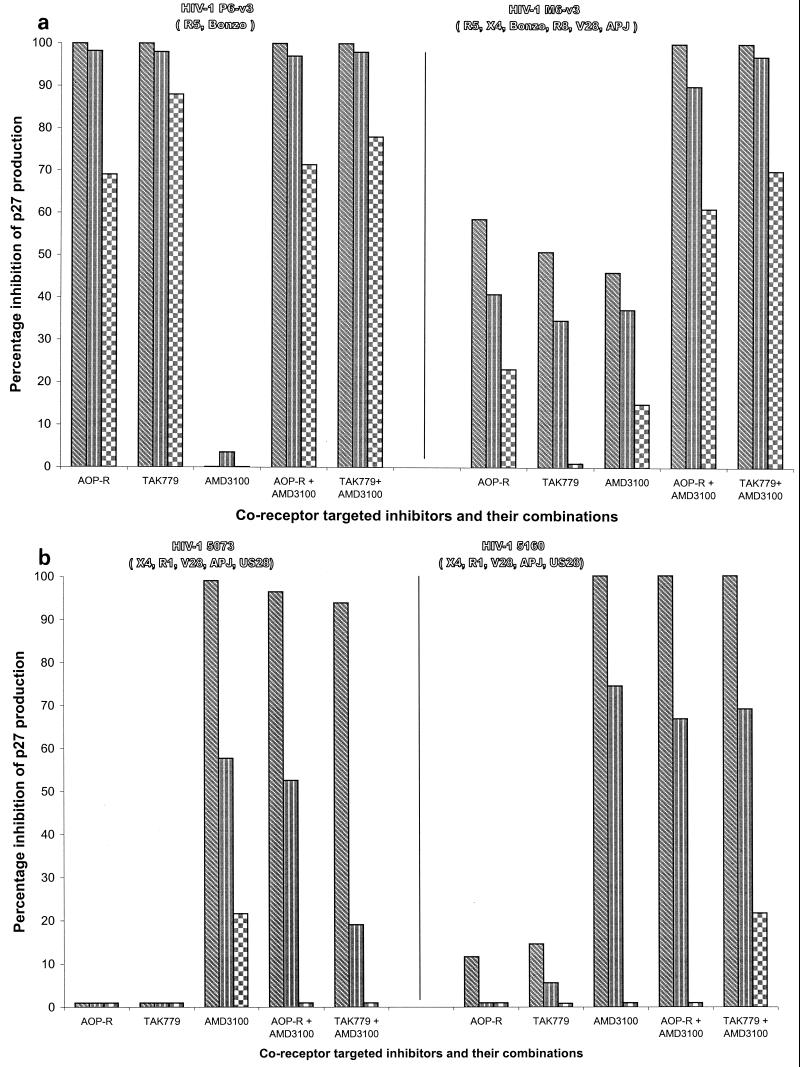
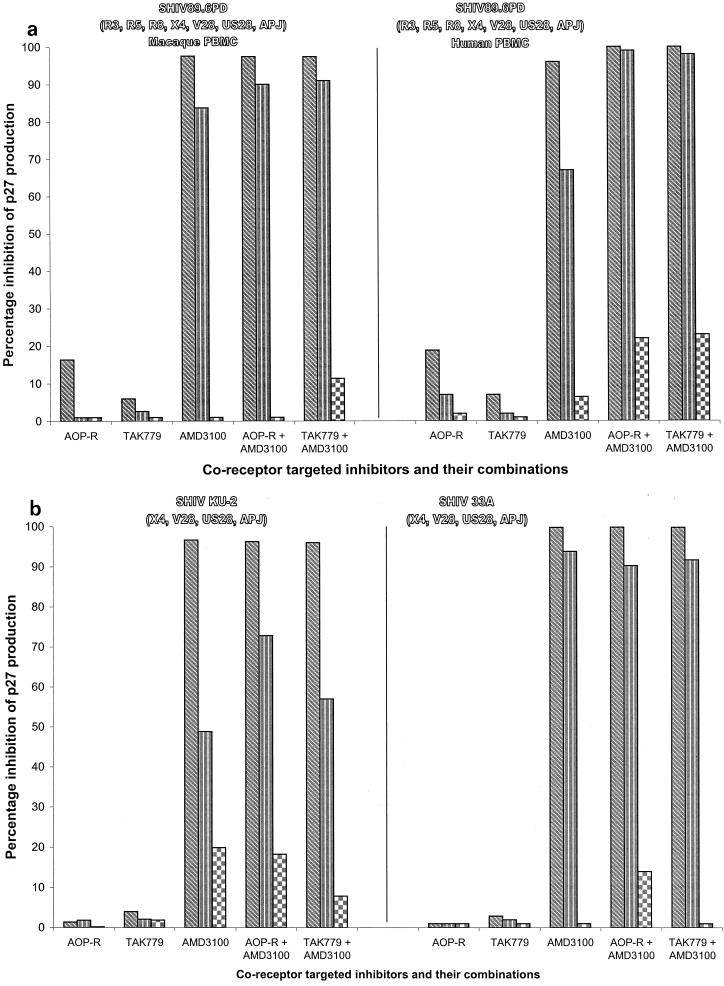
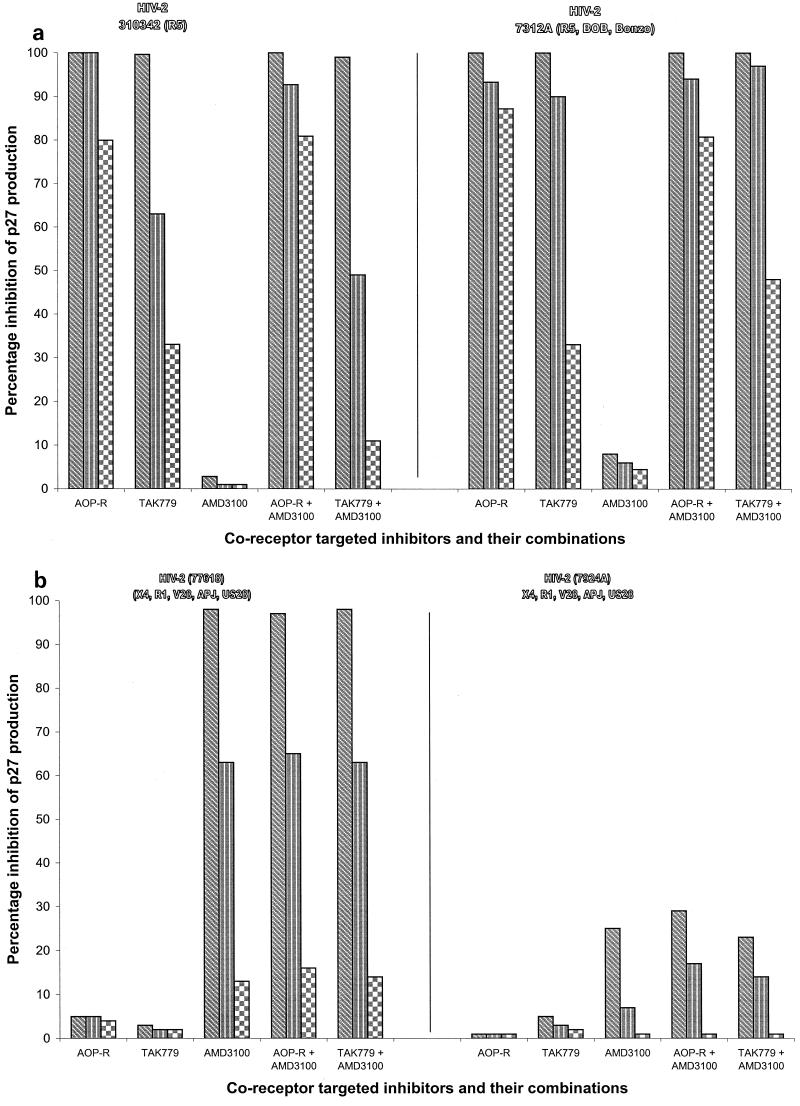
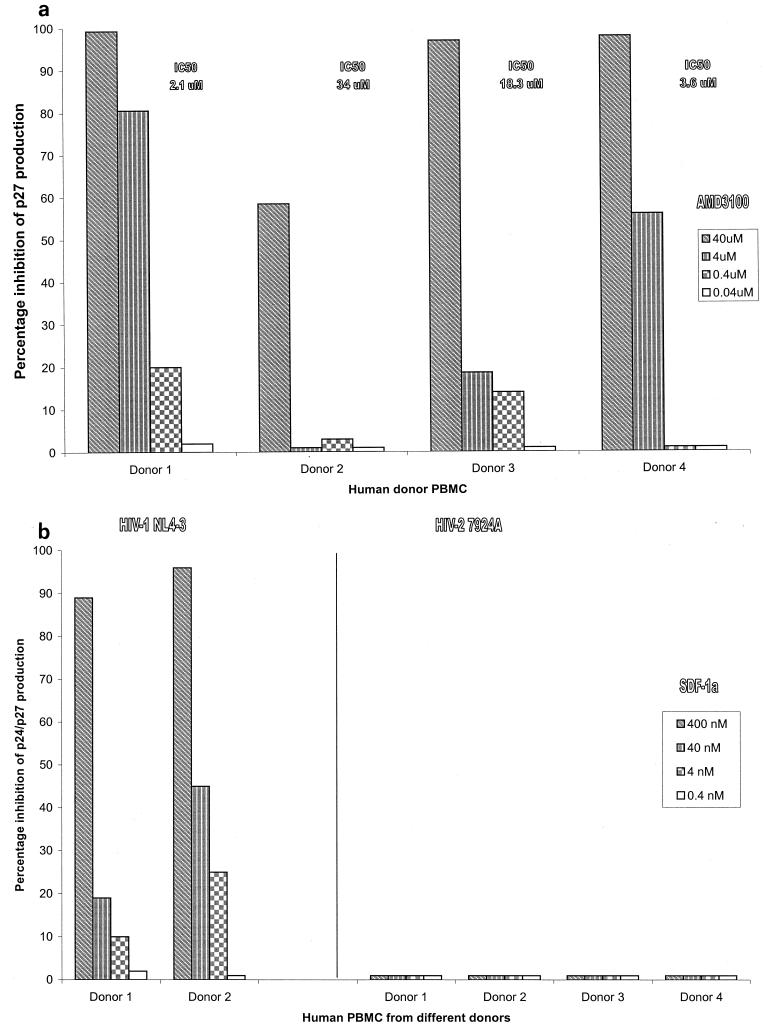
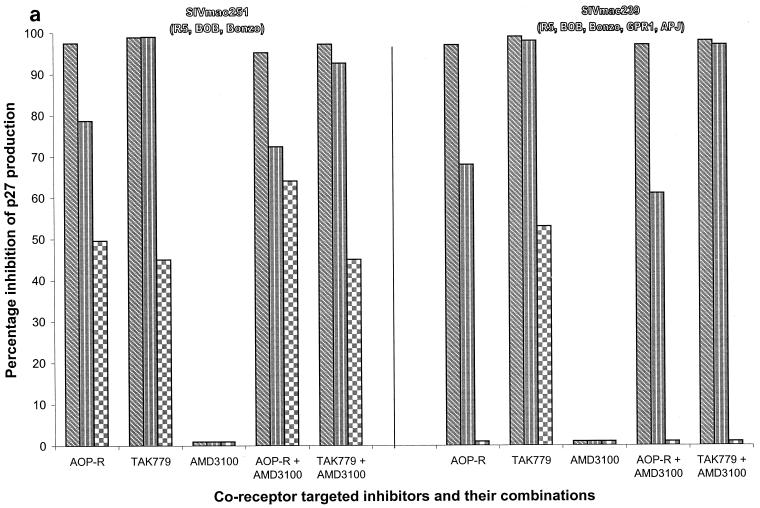
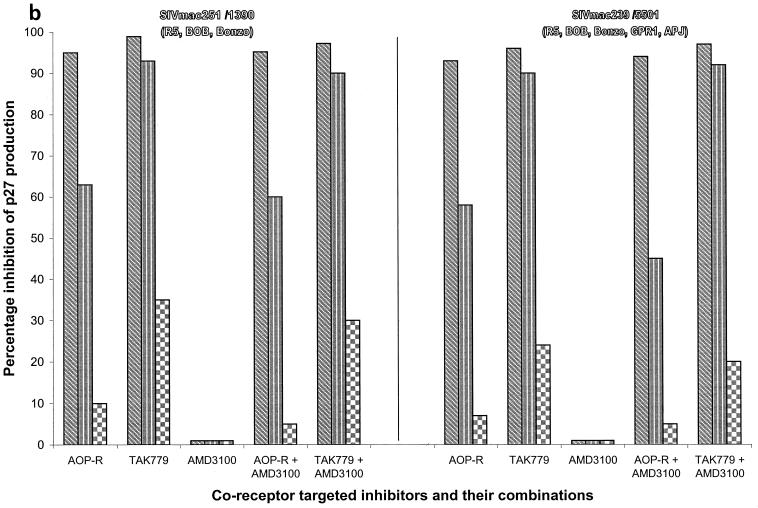
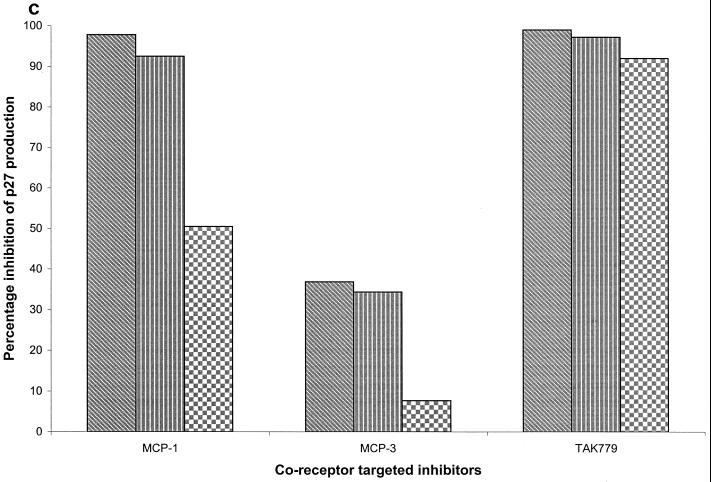
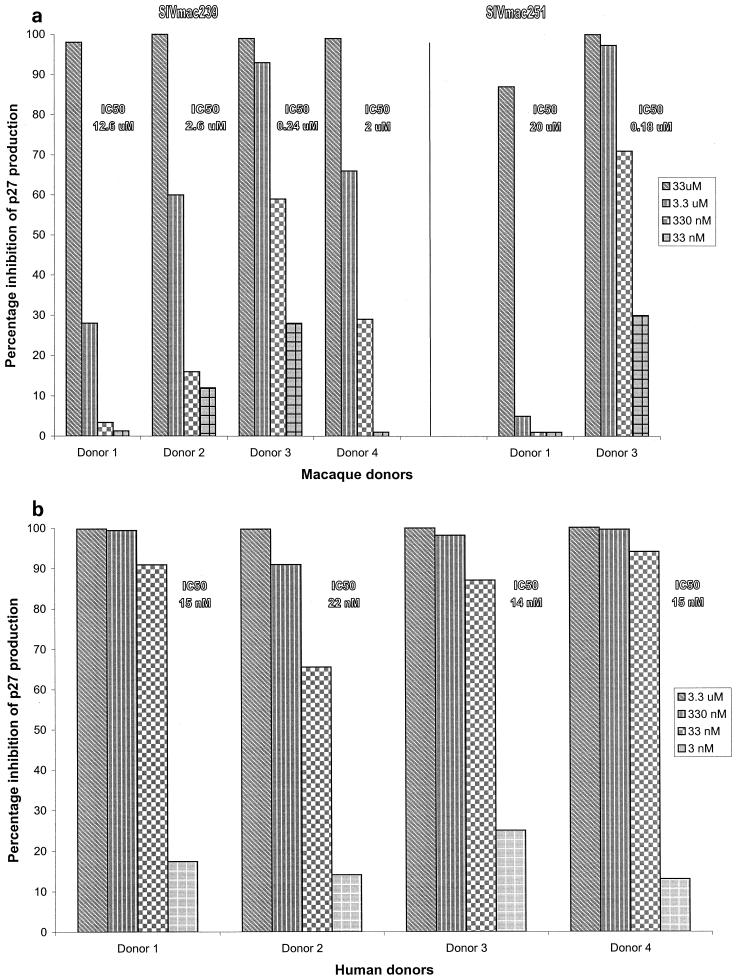
We have used coreceptor-targeted inhibitors to investigate which coreceptors are used by human immunodeficiency virus type 1 (HIV-1), simian immunodeficiency viruses (SIV), and human immunodeficiency virus type 2 (HIV-2) to enter peripheral blood mononuclear cells (PBMC). The inhibitors are TAK-779, which is specific for CCR5 and CCR2, aminooxypentane-RANTES, which blocks entry via CCR5 and CCR3, and AMD3100, which targets CXCR4. We found that for all the HIV-1 isolates and all but one of the HIV-2 isolates tested, the only relevant coreceptors were CCR5 and CXCR4. However, one HIV-2 isolate replicated in human PBMC even in the presence of TAK-779 and AMD3100, suggesting that it might use an undefined, alternative coreceptor that is expressed in the cells of some individuals. SIV(mac)239 and SIV(mac)251 (from macaques) were also able to use an alternative coreceptor to enter PBMC from some, but not all, human and macaque donors. The replication in human PBMC of SIV(rcm) (from a red-capped mangabey), a virus which uses CCR2 but not CCR5 for entry, was blocked by TAK-779, suggesting that CCR2 is indeed the paramount coreceptor for this virus in primary cells.
Figures
References
-
- Abebe A, Demissie D, Goudsmit J, Brouwer M, Kuiken C L, Pollakis G, Schuitemaker H, Fontanet A L, Rinke deWit T F. HIV-1 subtype C syncytium- and non-syncytium-inducing phenotypes and coreceptor usage among Ethiopian patients with AIDS. AIDS. 1999;13:1305–1311. - PubMed
-
- Alkhatib G, Combadiere C, Broder C C, Feng Y, Kennedy P E, Murphy P M, Berger E A. CC CKR5: a RANTES, MIP-1α, MIP-1β receptor as a fusion cofactor for macrophage-tropic HIV-1. Science. 1996;272:1955–1958. - PubMed
-
- Alkhatib G, Liao F, Berger E A, Farber J M, Peden K W C. A new SIV coreceptor, STRL33. Nature. 1997;388:238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
