

Cell-specific and conditional expression of caffeoyl-coenzyme A-3-O-methyltransferase in poplar
- PMID: 10889235
- PMCID: PMC59049
- DOI: 10.1104/pp.123.3.853
Cell-specific and conditional expression of caffeoyl-coenzyme A-3-O-methyltransferase in poplar
Abstract
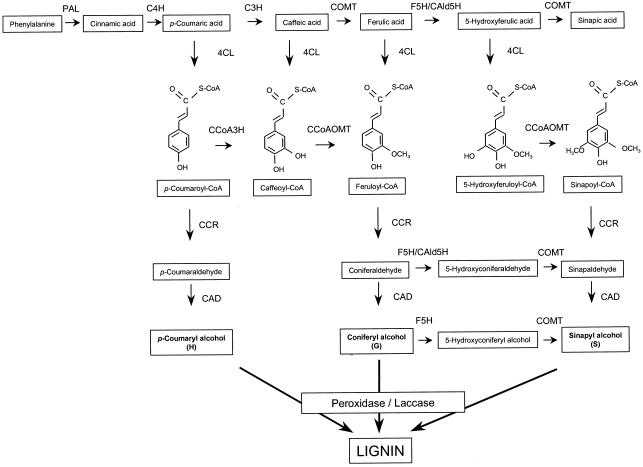
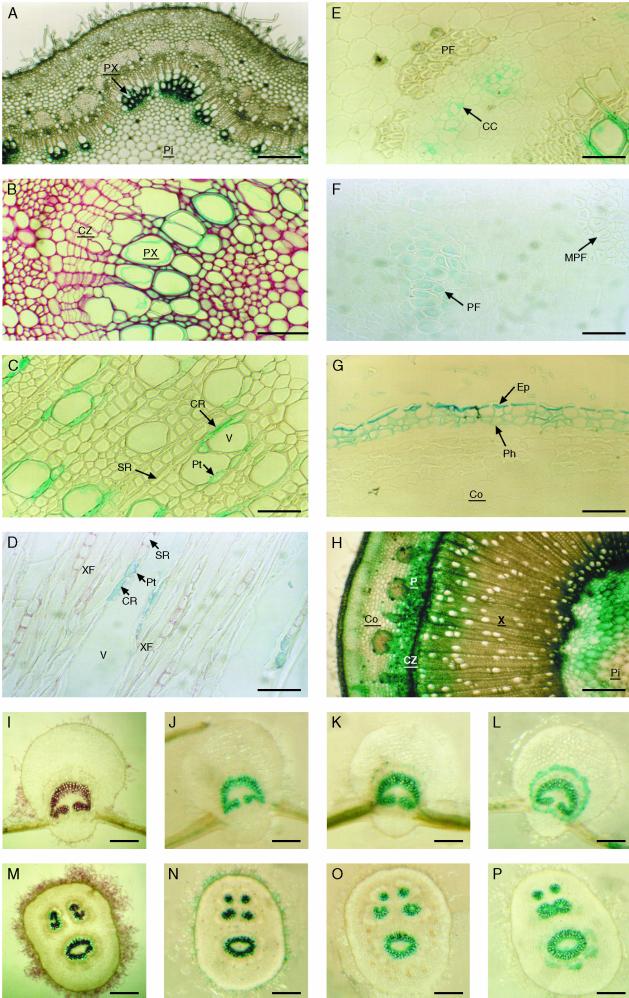
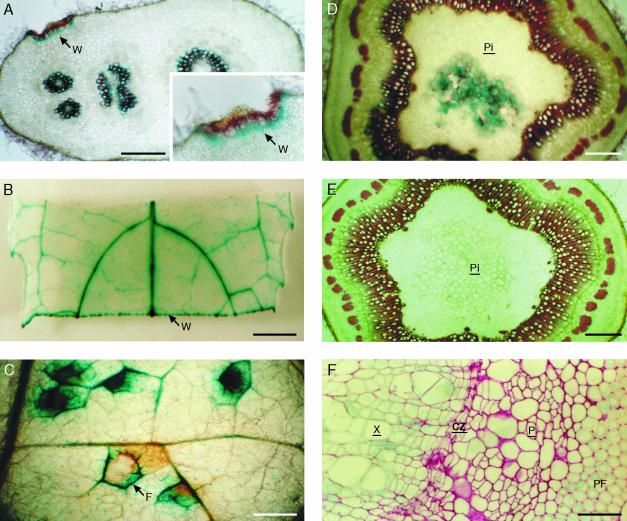
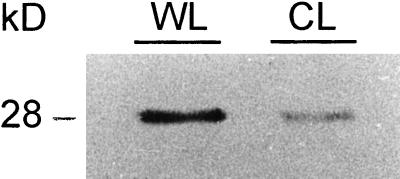
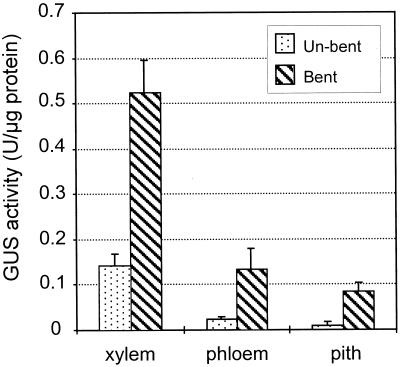
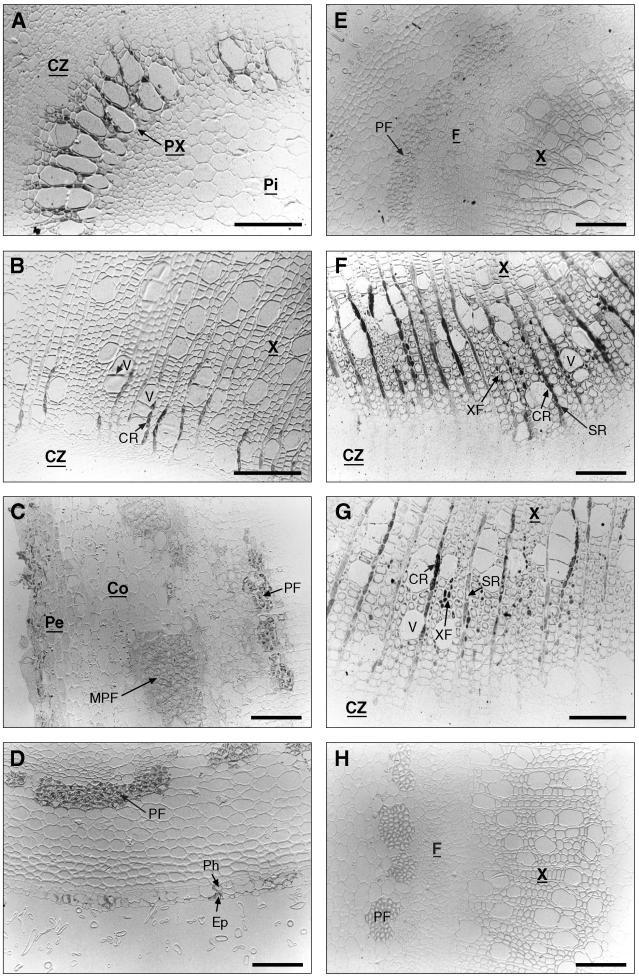
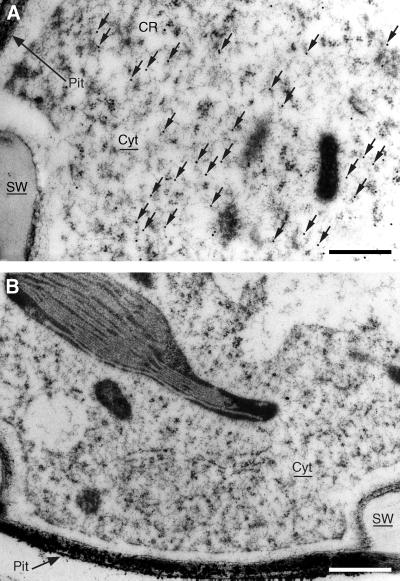
Caffeoyl coenzyme A-3-O-methyltransferase (CCoAOMT) plays an important role in lignin biosynthesis and is encoded by two genes in poplar (Populus trichocarpa). Here, we describe the expression pattern conferred by the two CCoAOMT promoters when fused to the gus-coding sequence in transgenic poplar (Populus tremula x Populus alba). Both genes were expressed similarly in xylem and differentially in phloem. In xylem, expression was preferentially observed in vessels and contact rays, whereas expression was barely detectable in storage rays and fibers, suggesting different routes to monolignol biosynthesis in the different xylem types. Furthermore, after wounding, fungal infection, and bending, the expression of both genes was induced concomitantly with de novo lignin deposition. Importantly, upon bending and leaning of the stem, the cell-specific expression pattern was lost, and both genes were expressed in all cell types of the xylem. CCoAOMT promoter activity correlated well with the presence of the CCoAOMT protein, as shown by immunolocalization. These expression data may explain, at least in part, the heterogeneity in lignin composition that is observed between cell types and upon different environmental conditions.
Figures
References
-
- Atanassova R, Favet N, Martz F, Chabbert B, Tollier MT, Monties B, Fritig B, Legrand M. Altered lignin composition in transgenic tobacco expressing O-methyltransferase sequences in sense and antisense orientation. Plant J. 1995;8:465–477.
-
- Baucher M, Monties B, Van Montagu M, Boerjan W. Biosynthesis and genetic engineering of lignin. Crit Rev Plant Sci. 1998;17:125–197.
-
- Blanchette RA, Biggs AR. Defense Mechanisms of Woody Plants against Fungi (Springer Series in Wood Science). Berlin: Springer-Verlag; 1992.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
