

Isolation and characterization of plant N-acetyl glucosaminyltransferase I (GntI) cDNA sequences. Functional analyses in the Arabidopsis cgl mutant and in antisense plants
- PMID: 10889259
- PMCID: PMC59073
- DOI: 10.1104/pp.123.3.1097
Isolation and characterization of plant N-acetyl glucosaminyltransferase I (GntI) cDNA sequences. Functional analyses in the Arabidopsis cgl mutant and in antisense plants
Abstract
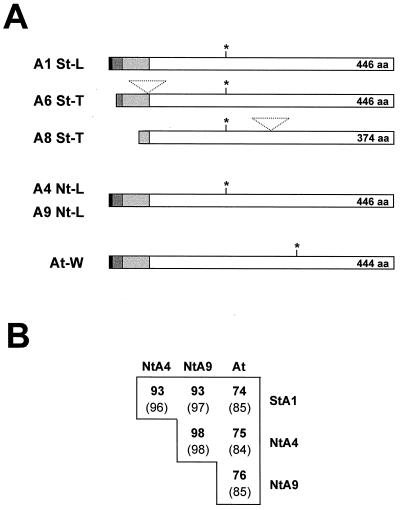
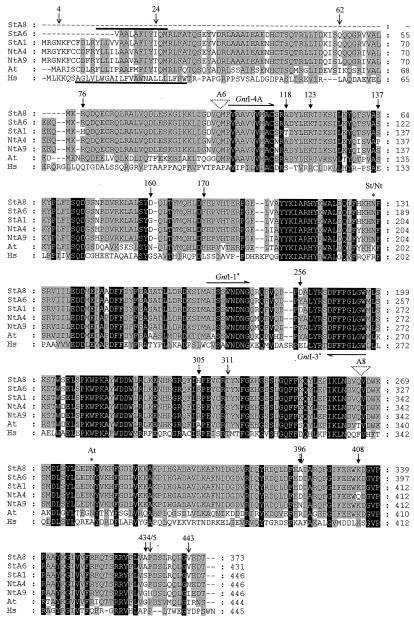
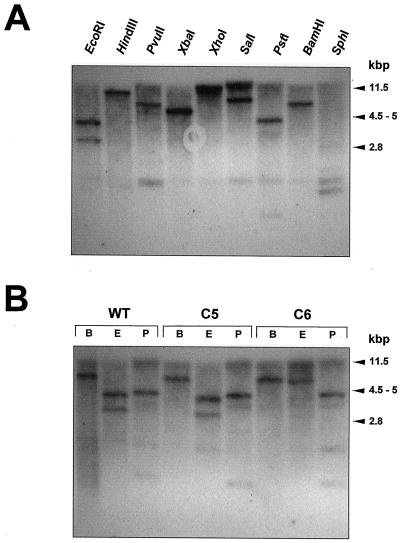
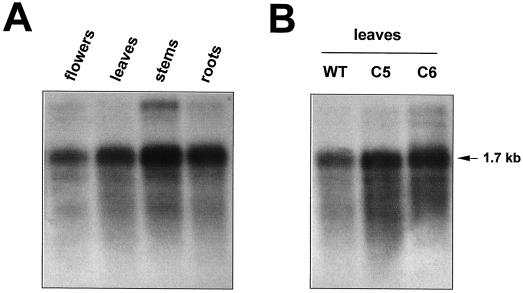
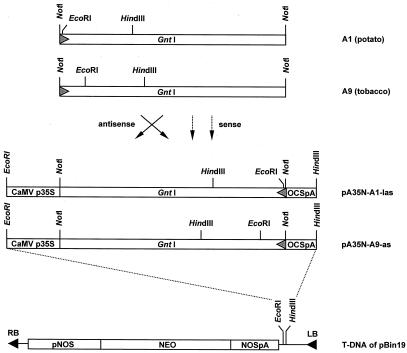
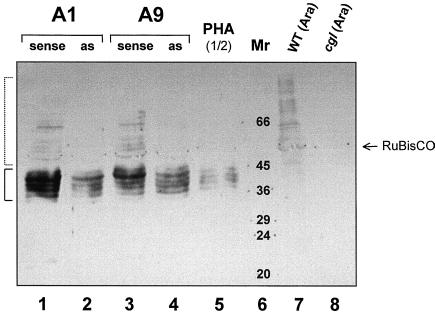
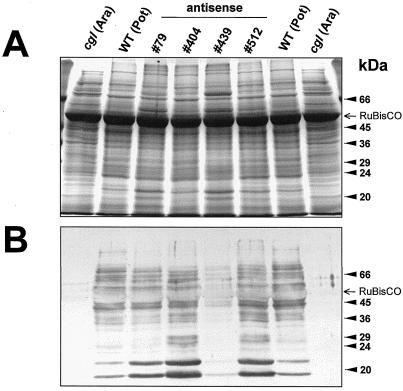
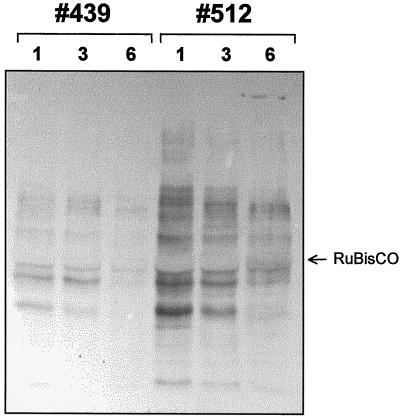
We report on the isolation and characterization of full-length cDNA sequences coding for N-acetylglucosaminyltransferase I (GnTI) from potato (Solanum tuberosum L.), tobacco (Nicotiana tabacum L.), and Arabidopsis. The deduced polypeptide sequences show highest homology among the solanaceous species (93% identity between potato and tobacco compared with about 75% with Arabidopsis) but share only weak homology with human GnTI (35% identity). In contrast to the corresponding enzymes from animals, all plant GnTI sequences identified are characterized by a much shorter hydrophobic membrane anchor and contain one putative N-glycosylation site that is conserved in potato and tobacco, but differs in Arabidopsis. Southern-blot analyses revealed that GntI behaves as a single-copy gene. Northern-blot analyses showed that GntI-mRNA expression is largely constitutive. Arabidopsis cgl mutants deficient in GnTI activity also possess GntI mRNA, indicating that they result from point mutations. GntI-expression constructs were tested for the ability to relieve the GnTI block in protoplasts of the Arabidopsis cgl mutant and used to obtain transgenic potato and tobacco plants that display a substantial reduction of complex glycan patterns. The latter observation indicates that production of heterologous glycoproteins with little or no antigenic glycans can be achieved in whole plants, and not in just Arabidopsis, using antisense technology.
Figures
Similar articles
-
Molecular basis of N-acetylglucosaminyltransferase I deficiency in Arabidopsis thaliana plants lacking complex N-glycans.Biochem J. 2005 Apr 15;387(Pt 2):385-91. doi: 10.1042/BJ20041686. Biochem J. 2005. PMID: 15537386 Free PMC article.
-
Unaltered complex N-glycan profiles in Nicotiana benthamiana despite drastic reduction of beta1,2- N -acetylglucosaminyltransferase I activity.Glycoconj J. 2004;21(5):275-82. doi: 10.1023/B:GLYC.0000045099.29038.04. Glycoconj J. 2004. PMID: 15486460
-
Molecular cloning and characterization of cDNA coding for beta1, 2N-acetylglucosaminyltransferase I (GlcNAc-TI) from Nicotiana tabacum.Glycobiology. 1999 Aug;9(8):779-85. doi: 10.1093/glycob/9.8.779. Glycobiology. 1999. PMID: 10406843
-
Tissues of the clawed frog Xenopus laevis contain two closely related forms of UDP-GlcNAc:alpha3-D-mannoside beta-1,2-N-acetylglucosaminyltransferase I.Glycobiology. 2001 Sep;11(9):769-78. doi: 10.1093/glycob/11.9.769. Glycobiology. 2001. PMID: 11555621
-
Assessment of technological options and economical feasibility for cyanophycin biopolymer and high-value amino acid production.Appl Microbiol Biotechnol. 2007 Nov;77(2):257-67. doi: 10.1007/s00253-007-1178-3. Epub 2007 Sep 18. Appl Microbiol Biotechnol. 2007. PMID: 17876577 Free PMC article. Review.
Cited by
-
Reduction of cross-reactive carbohydrate determinants in plant foodstuff: elucidation of clinical relevance and implications for allergy diagnosis.PLoS One. 2011 Mar 14;6(3):e17800. doi: 10.1371/journal.pone.0017800. PLoS One. 2011. PMID: 21423762 Free PMC article.
-
Cell wall O-glycoproteins and N-glycoproteins: aspects of biosynthesis and function.Front Plant Sci. 2014 Oct 2;5:499. doi: 10.3389/fpls.2014.00499. eCollection 2014. Front Plant Sci. 2014. PMID: 25324850 Free PMC article. Review.
-
Comparative analyses of Arabidopsis complex glycan1 mutants and genetic interaction with staurosporin and temperature sensitive3a.Plant Physiol. 2008 Nov;148(3):1354-67. doi: 10.1104/pp.108.127027. Epub 2008 Sep 3. Plant Physiol. 2008. PMID: 18768906 Free PMC article.
-
Recent Developments in Deciphering the Biological Role of Plant Complex N-Glycans.Front Plant Sci. 2022 Apr 25;13:897549. doi: 10.3389/fpls.2022.897549. eCollection 2022. Front Plant Sci. 2022. PMID: 35557740 Free PMC article. Review.
-
OsMOGS is required for N-glycan formation and auxin-mediated root development in rice (Oryza sativa L.).Plant J. 2014 May;78(4):632-645. doi: 10.1111/tpj.12497. Epub 2014 Apr 15. Plant J. 2014. PMID: 24597623 Free PMC article.
References
-
- Boer GJ, Testerink C, Pielage G, Nijkamp HJ, Stuitje AR. Sequences surrounding the transcription initiation site in the Arabidopsis enoyl-acyl carrier protein reductase gene control seed expression in transgenic tobacco. Plant Mol Biol. 1999;39:1197–1207. - PubMed
-
- Dahms NM, Lobel P, Kornfeld S. Mannose-6-phosphate receptors and lysosomal enzyme targeting. J Biol Chem. 1989;264:12115–12118. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
