

Enhancement of induced disease resistance by simultaneous activation of salicylate- and jasmonate-dependent defense pathways in Arabidopsis thaliana
- PMID: 10890883
- PMCID: PMC27013
- DOI: 10.1073/pnas.130425197
Enhancement of induced disease resistance by simultaneous activation of salicylate- and jasmonate-dependent defense pathways in Arabidopsis thaliana
Abstract
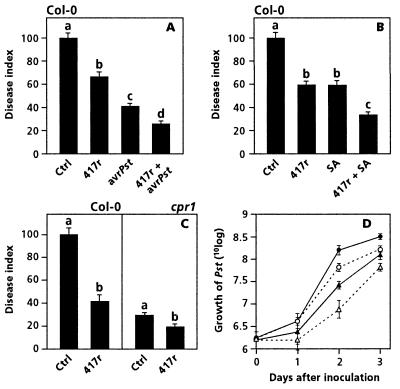
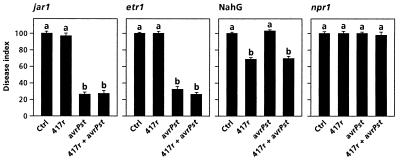
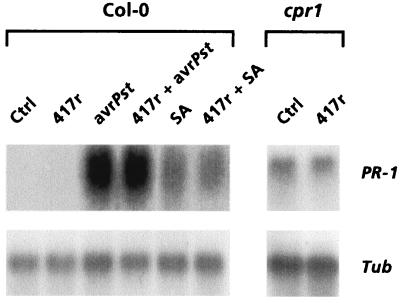
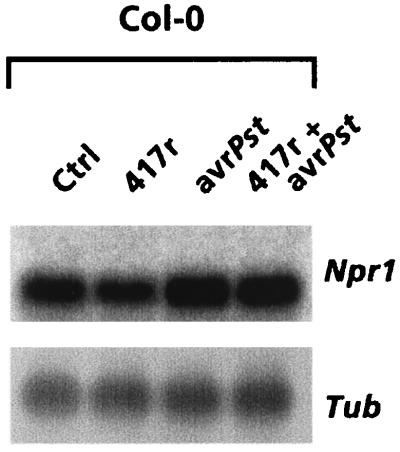
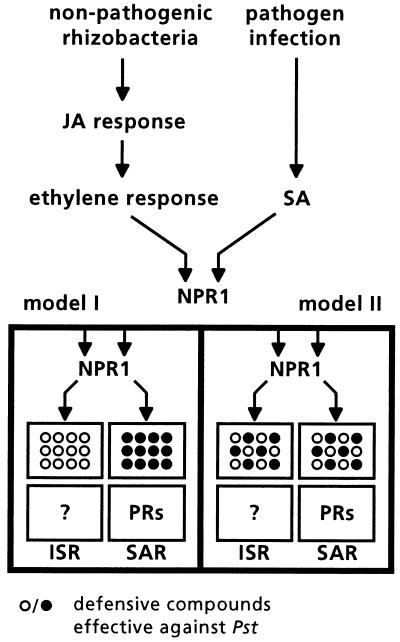
The plant-signaling molecules salicylic acid (SA) and jasmonic acid (JA) play an important role in induced disease resistance pathways. Cross-talk between SA- and JA-dependent pathways can result in inhibition of JA-mediated defense responses. We investigated possible antagonistic interactions between the SA-dependent systemic acquired resistance (SAR) pathway, which is induced upon pathogen infection, and the JA-dependent induced systemic resistance (ISR) pathway, which is triggered by nonpathogenic Pseudomonas rhizobacteria. In Arabidopsis thaliana, SAR and ISR are effective against a broad spectrum of pathogens, including the foliar pathogen Pseudomonas syringae pv. tomato (Pst). Simultaneous activation of SAR and ISR resulted in an additive effect on the level of induced protection against Pst. In Arabidopsis genotypes that are blocked in either SAR or ISR, this additive effect was not evident. Moreover, induction of ISR did not affect the expression of the SAR marker gene PR-1 in plants expressing SAR. Together, these observations demonstrate that the SAR and the ISR pathway are compatible and that there is no significant cross-talk between these pathways. SAR and ISR both require the key regulatory protein NPR1. Plants expressing both types of induced resistance did not show elevated Npr1 transcript levels, indicating that the constitutive level of NPR1 is sufficient to facilitate simultaneous expression of SAR and ISR. These results suggest that the enhanced level of protection is established through parallel activation of complementary, NPR1-dependent defense responses that are both active against Pst. Therefore, combining SAR and ISR provides an attractive tool for the improvement of disease control.
Figures
References
-
- Reymond P, Farmer E E. Curr Opin Plant Biol. 1998;1:404–411. - PubMed
-
- Bostock R M. Physiol Mol Plant Pathol. 1999;55:99–109.
-
- Glazebrook J. Curr Opin Plant Biol. 1999;2:280–286. - PubMed
-
- Maleck K, Dietrich R A. Trends Plant Sci. 1999;4:215–219. - PubMed
-
- Pieterse C M J, Van Loon L C. Trends Plant Sci. 1999;4:52–58. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
