

Phosphate is a specific signal for induction of osteopontin gene expression
- PMID: 10890885
- PMCID: PMC26951
- DOI: 10.1073/pnas.140021997
Phosphate is a specific signal for induction of osteopontin gene expression
Abstract
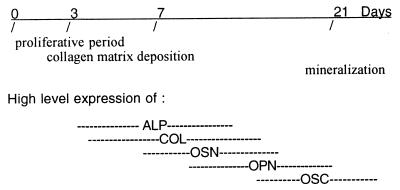
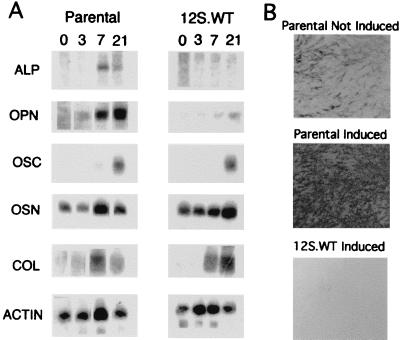
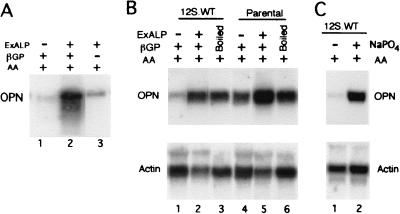
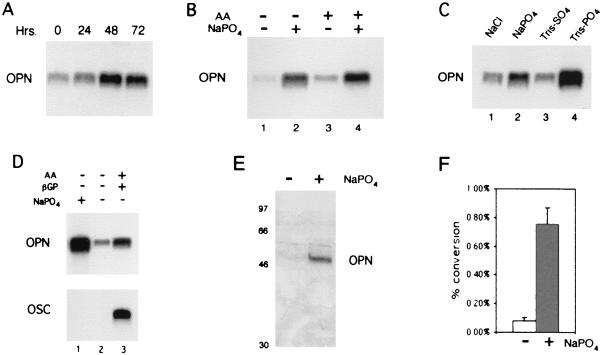
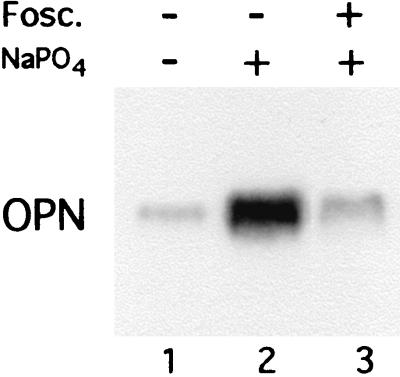
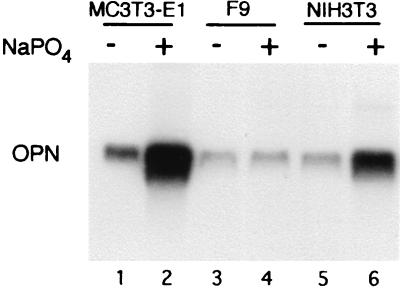
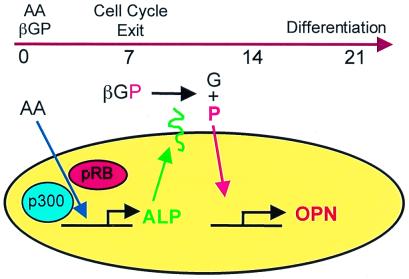
Osteopontin is a phosphorylated glycoprotein secreted to the mineralizing extracellular matrix by osteoblasts during bone development. It is believed to facilitate the attachment of osteoblasts and osteoclasts to the extracellular matrix, allowing them to perform their respective functions during osteogenesis. Several other functions have been suggested for this protein, and its up-regulation is associated with various disease states related to calcification, including arterial plaque formation and the formation of kidney stones. Although expression of this gene has been demonstrated in multiple tissues, its regulation is not well understood. Our previous studies on the roles of the retinoblastoma protein (pRB) and p300/CBP in the regulation of osteoblast differentiation revealed a link between osteopontin induction and the synthesis of alkaline phosphatase. In this paper, we describe results specifically linking induction of osteopontin to the enzymatic activity of alkaline phosphatase in the medium, which results in the generation of free phosphate. This elevation of free phosphate in the medium is sufficient to signal induction of osteopontin RNA and protein. The strong and specific induction of osteopontin in direct response to increased phosphate levels provides a mechanism to explain how expression of this product is normally regulated in bone and suggests how it may become up-regulated in damaged tissue.
Figures
References
-
- Moran E. Semin Virol. 1994;5:327–340.
-
- Beck G R, Jr, Zerler B, Moran E. In: Human Tumor Viruses. McCance D J, editor. Washington, DC: Am. Soc. Microbiol.; 1998. pp. 51–86.
-
- Mayol X, Grana X. Prog Cell Cycle Res. 1997;3:157–169. - PubMed
-
- Giordano A, Avantiagatti M L. J Cell Physiol. 1999;181:218–230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
