

Bacterial-type DNA holliday junction resolvases in eukaryotic viruses
- PMID: 10890916
- PMCID: PMC16798
- DOI: 10.1073/pnas.150238697
Bacterial-type DNA holliday junction resolvases in eukaryotic viruses
Abstract
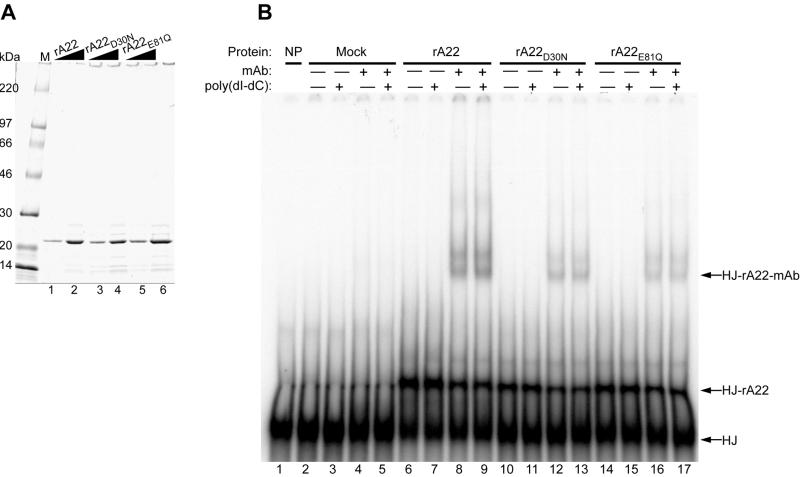
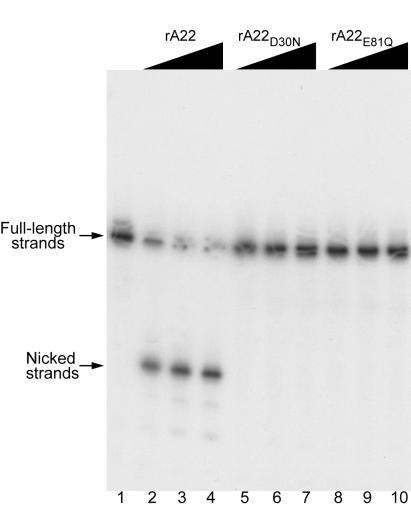
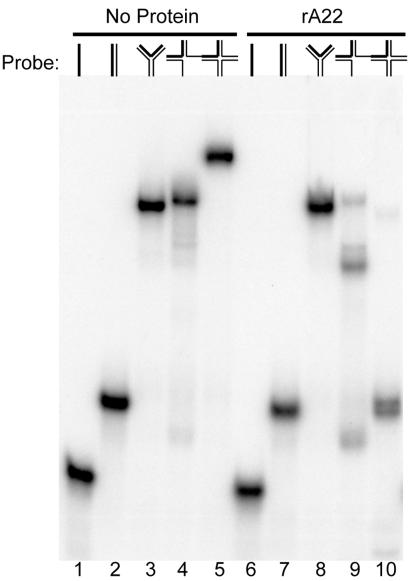
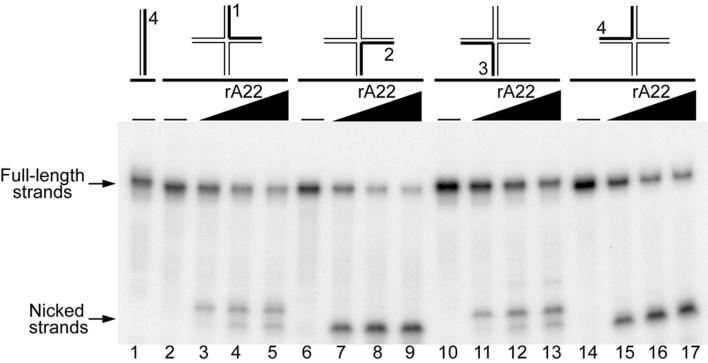
Homologous DNA recombination promotes genetic diversity and the maintenance of genome integrity, yet no enzymes with specificity for the Holliday junction (HJ)-a key DNA recombination intermediate-have been purified and characterized from metazoa or their viruses. Here we identify critical structural elements of RuvC, a bacterial HJ resolvase, in uncharacterized open reading frames from poxviruses and an iridovirus. The putative vaccinia virus resolvase was expressed as a recombinant protein, affinity purified, and shown to specifically bind and cleave a synthetic HJ to yield nicked duplex molecules. Mutation of either of two conserved acidic amino acids abrogated the catalytic activity of the A22R protein without affecting HJ binding. The presence of bacterial-type enzymes in metazoan viruses raises evolutionary questions.
Figures

Comment in
-
Resolving the relationships of resolving enzymes.Proc Natl Acad Sci U S A. 2000 Aug 15;97(17):9351-3. doi: 10.1073/pnas.97.17.9351. Proc Natl Acad Sci U S A. 2000. PMID: 10944205 Free PMC article. No abstract available.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
