

Dopamine D1 and adenosine A1 receptors form functionally interacting heteromeric complexes
- PMID: 10890919
- PMCID: PMC26995
- DOI: 10.1073/pnas.150241097
Dopamine D1 and adenosine A1 receptors form functionally interacting heteromeric complexes
Abstract
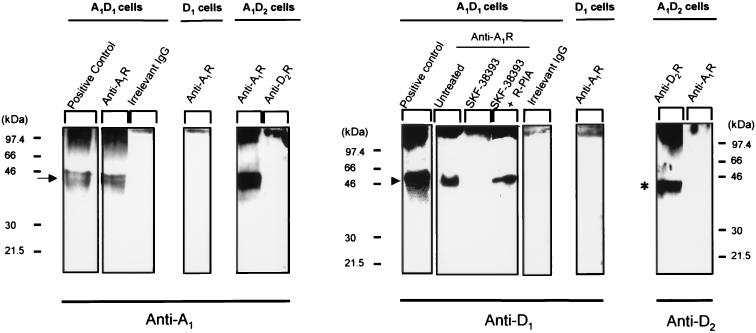
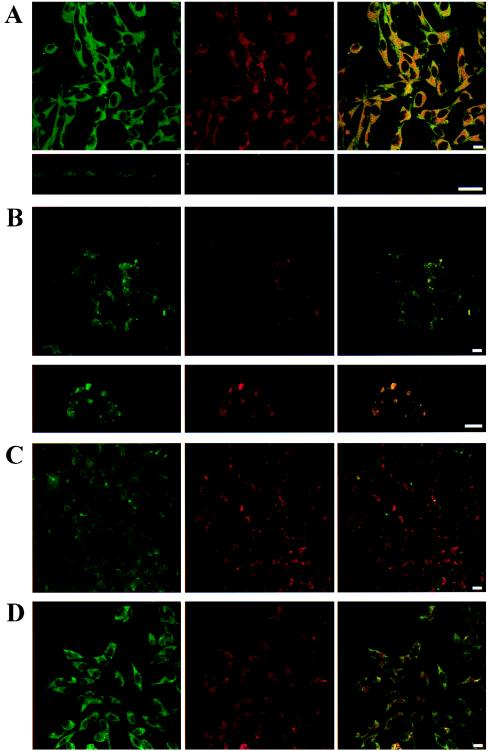
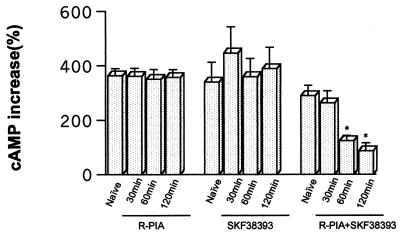
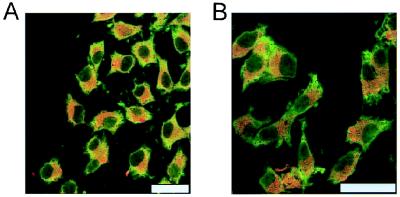
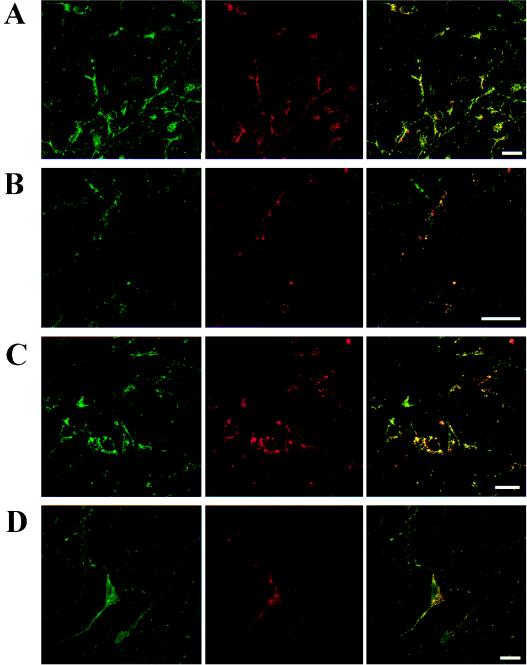
The possible molecular basis for the previously described antagonistic interactions between adenosine A(1) receptors (A(1)R) and dopamine D(1) receptors (D(1)R) in the brain have been studied in mouse fibroblast Ltk(-) cells cotransfected with human A(1)R and D(1)R cDNAs or with human A(1)R and dopamine D(2) receptor (long-form) (D(2)R) cDNAs and in cortical neurons in culture. A(1)R and D(1)R, but not A(1)R and D(2)R, were found to coimmunoprecipitate in cotransfected fibroblasts. This selective A(1)R/D(1)R heteromerization disappeared after pretreatment with the D(1)R agonist, but not after combined pretreatment with D(1)R and A(1)R agonists. A high degree of A(1)R and D(1)R colocalization, demonstrated in double immunofluorescence experiments with confocal laser microscopy, was found in both cotransfected fibroblast cells and cortical neurons in culture. On the other hand, a low degree of A(1)R and D(2)R colocalization was observed in cotransfected fibroblasts. Pretreatment with the A(1)R agonist caused coclustering (coaggregation) of A(1)R and D(1)R, which was blocked by combined pretreatment with the D(1)R and A(1)R agonists in both fibroblast cells and in cortical neurons in culture. Combined pretreatment with D(1)R and A(1)R agonists, but not with either one alone, substantially reduced the D(1)R agonist-induced accumulation of cAMP. The A(1)R/D(1)R heteromerization may be one molecular basis for the demonstrated antagonistic modulation of A(1)R of D(1)R receptor signaling in the brain. The persistence of A(1)R/D(1)R heteromerization seems to be essential for the blockade of A(1)R agonist-induced A(1)R/D(1)R coclustering and for the desensitization of the D(1)R agonist-induced cAMP accumulation seen on combined pretreatment with D(1)R and A(1)R agonists, which indicates a potential role of A(1)R/D(1)R heteromers also in desensitization mechanisms and receptor trafficking.
Figures
References
-
- Agnati L F, Fuxe K, Benfenati F, Celani M F, Battistini N, Mutt V, Cavicchioli L, Galli G, Hokfelt T. Neurosci Lett. 1983;35:179–183. - PubMed
-
- Fuxe K, Agnati L F, editors. Receptor-Receptor Interactions. Wenner-Gren Center Interactional Symposium Series. New York: Macmillan; 1987.
-
- Zoli M, Agnati L F, Hedlund P B, Li X M, Ferré S, Fuxe K. Mol Neurobiol. 1993;7:293–334. - PubMed
-
- Ng G Y, George S R, Zastawny R L, Caron M, Bouvier M, Dennis M, O'Dowd B F. Biochemistry. 1993;32:11727–11733. - PubMed
-
- Ng G Y, O'Dowd B F, Caron M, Dennis M, Brann M R, George S R. J Neurochem. 1994;63:1589–1595. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
