

Bop1 is a mouse WD40 repeat nucleolar protein involved in 28S and 5. 8S RRNA processing and 60S ribosome biogenesis
- PMID: 10891491
- PMCID: PMC86002
- DOI: 10.1128/MCB.20.15.5516-5528.2000
Bop1 is a mouse WD40 repeat nucleolar protein involved in 28S and 5. 8S RRNA processing and 60S ribosome biogenesis
Abstract
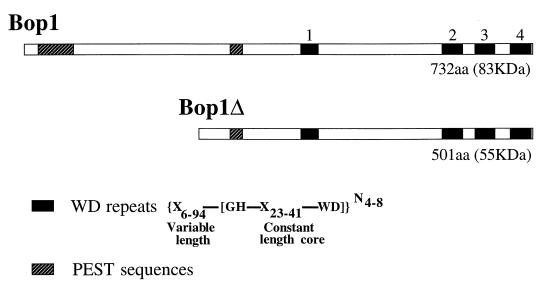
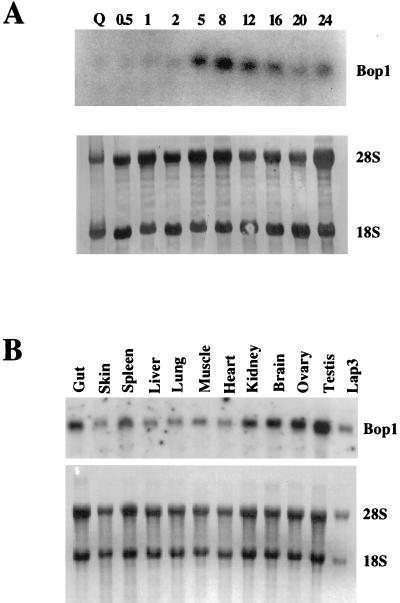
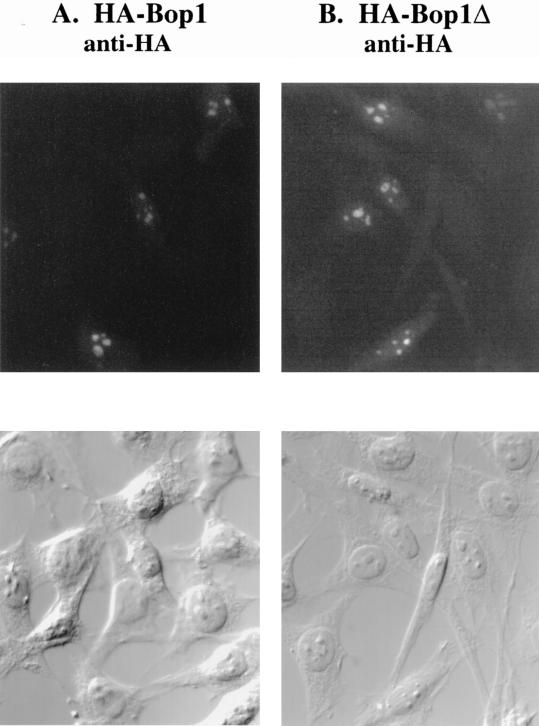
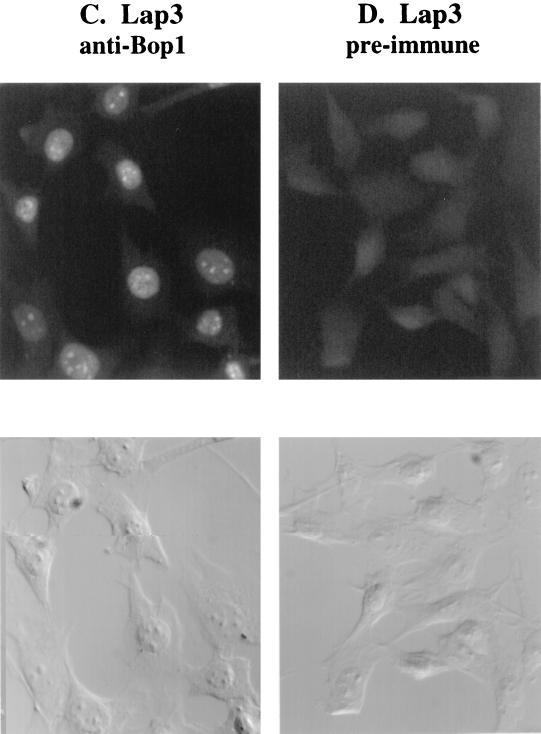
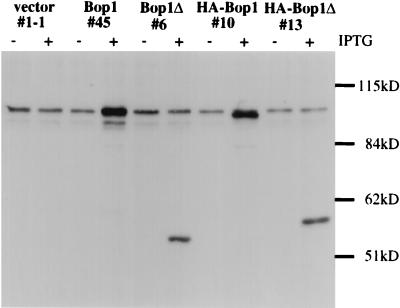
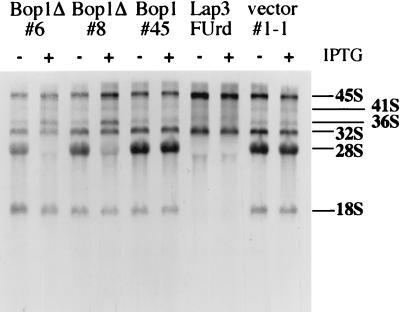
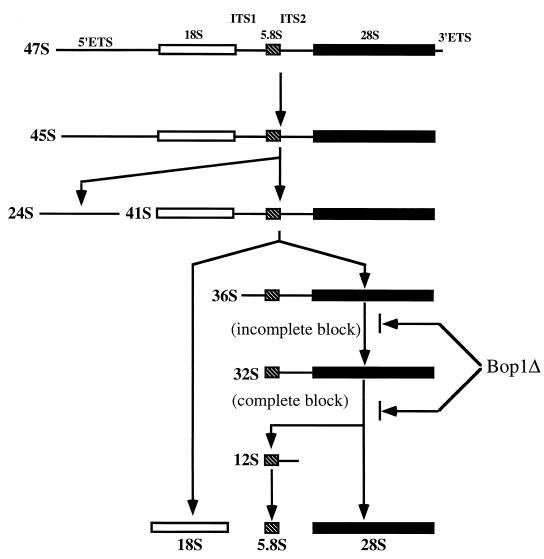
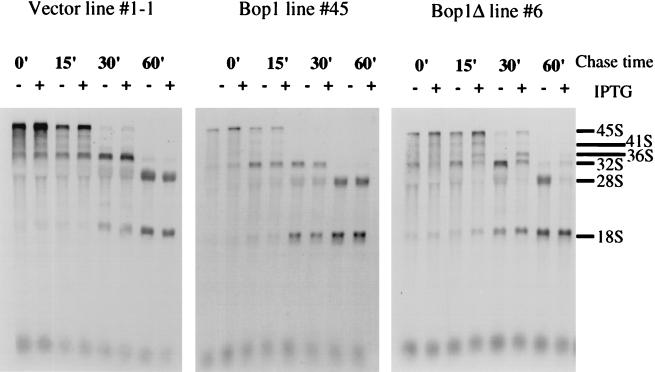
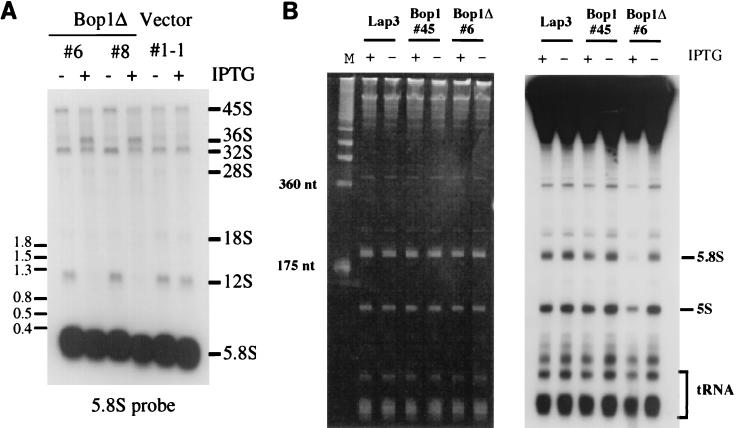
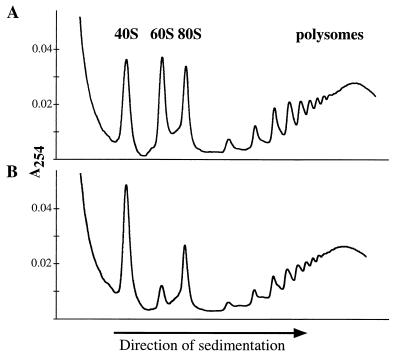
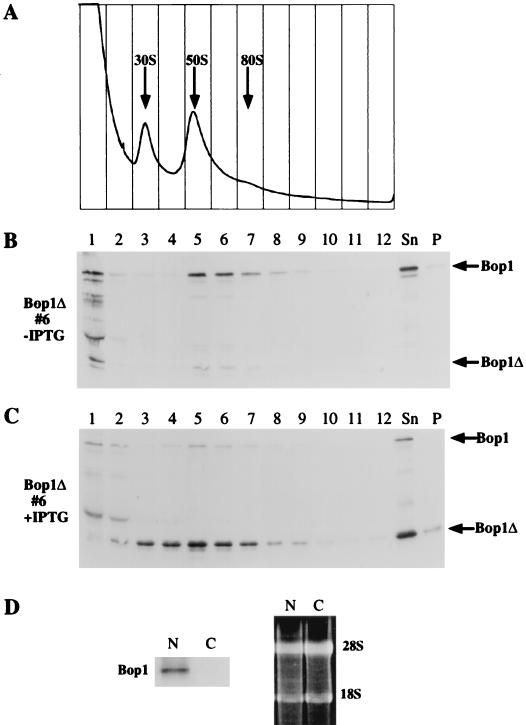
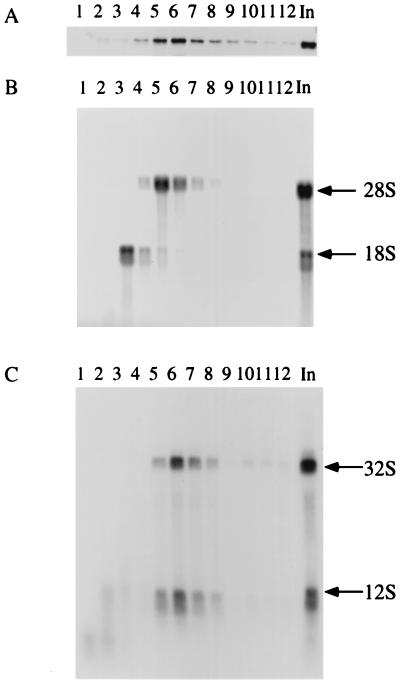
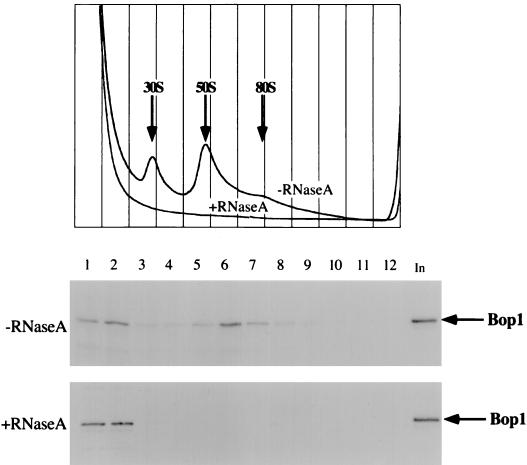
We have identified and characterized a novel mouse protein, Bop1, which contains WD40 repeats and is highly conserved through evolution. bop1 is ubiquitously expressed in all mouse tissues examined and is upregulated during mid-G(1) in serum-stimulated fibroblasts. Immunofluorescence analysis shows that Bop1 is localized predominantly to the nucleolus. In sucrose density gradients, Bop1 from nuclear extracts cosediments with the 50S-80S ribonucleoprotein particles that contain the 32S rRNA precursor. RNase A treatment disrupts these particles and releases Bop1 into a low-molecular-weight fraction. A mutant form of Bop1, Bop1Delta, which lacks 231 amino acids in the N- terminus, is colocalized with wild-type Bop1 in the nucleolus and in ribonucleoprotein complexes. Expression of Bop1Delta leads to cell growth arrest in the G(1) phase and results in a specific inhibition of the synthesis of the 28S and 5.8S rRNAs without affecting 18S rRNA formation. Pulse-chase analyses show that Bop1Delta expression results in a partial inhibition in the conversion of the 36S to the 32S pre-rRNA and a complete inhibition of the processing of the 32S pre-rRNA to form the mature 28S and 5.8S rRNAs. Concomitant with these defects in rRNA processing, expression of Bop1Delta in mouse cells leads to a deficit in the cytosolic 60S ribosomal subunits. These studies thus identify Bop1 as a novel, nonribosomal mammalian protein that plays a key role in the formation of the mature 28S and 5.8S rRNAs and in the biogenesis of the 60S ribosomal subunit.
Figures
References
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1994.
-
- Bachant J B, Elledge S J. Mitotic treasures in the nucleolus. Nature. 1999;398:757–758. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases