

beta-1,2-linked oligomannosides from Candida albicans bind to a 32-kilodalton macrophage membrane protein homologous to the mammalian lectin galectin-3
- PMID: 10899835
- PMCID: PMC98331
- DOI: 10.1128/IAI.68.8.4391-4398.2000
beta-1,2-linked oligomannosides from Candida albicans bind to a 32-kilodalton macrophage membrane protein homologous to the mammalian lectin galectin-3
Abstract
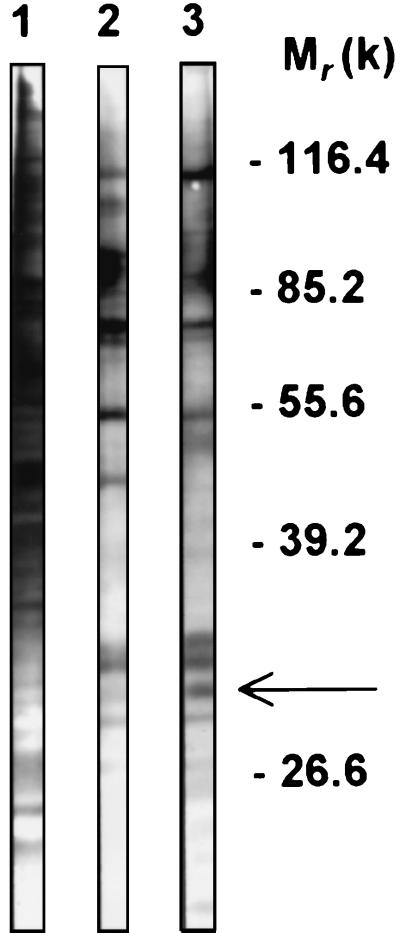
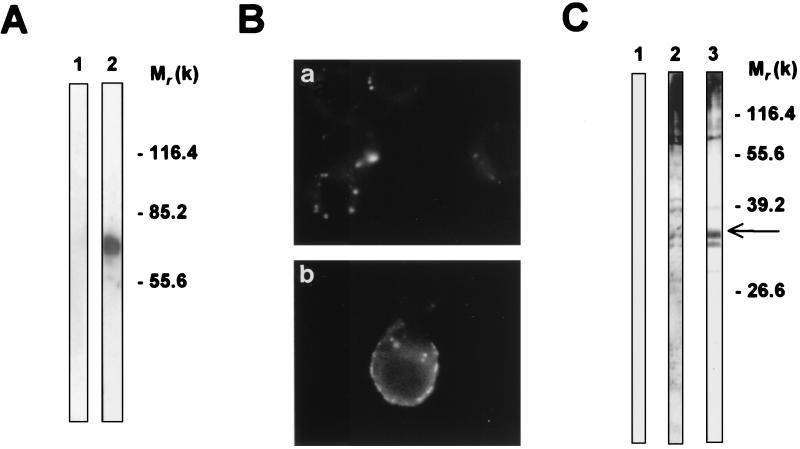
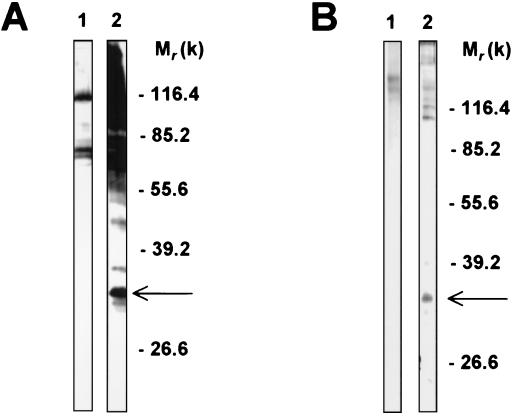
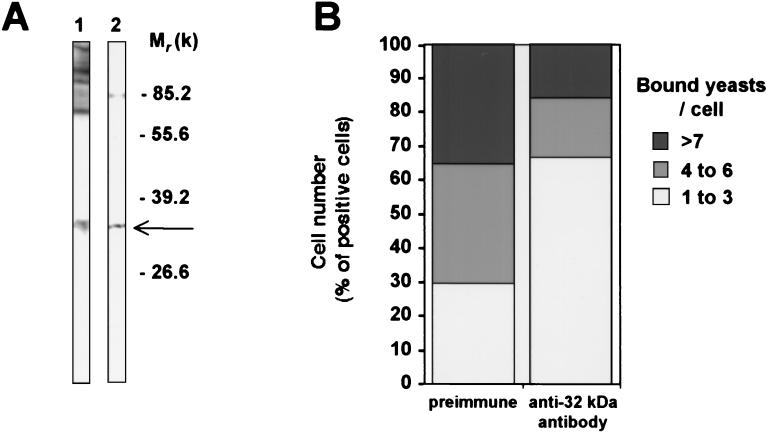
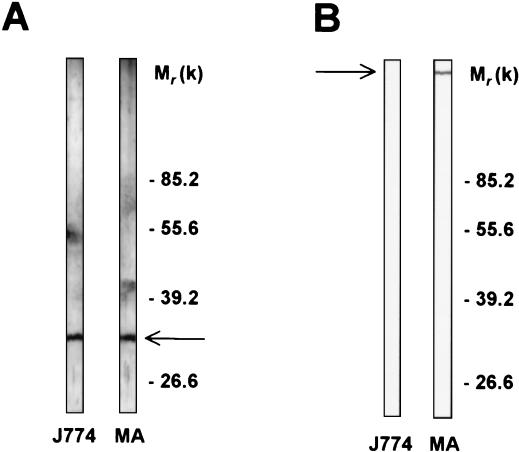
beta-1,2-linked oligomannoside residues are present, associated with mannan and a glycolipid, the phospholipomannan, at the Candida albicans cell wall surface. beta-1,2-linked oligomannoside residues act as adhesins for macrophages and stimulate these cells to undergo cytokine production. To characterize the macrophage receptor involved in the recognition of C. albicans beta-1,2-oligomannoside we used the J774 mouse cell line, which is devoid of the receptor specific for alpha-linked mannose residues. A series of experiments based on affinity binding on either C. albicans yeast cells or beta-1,2-oligomannoside-conjugated bovine serum albumin (BSA) and subsequent disclosure with biotinylated conjugated BSA repeatedly led to the detection of a 32-kDa macrophage protein. An antiserum specific for this 32-kDa protein inhibited C. albicans binding to macrophages and was used to immunoprecipitate the molecule. Two high-pressure liquid chromatography-purified peptides from the 32-kDa tryptic digest showed complete homology to galectin-3 (previously designated Mac-2 antigen), an endogenous lectin with pleiotropic functions which is expressed in a wide variety of cell types with which C. albicans interacts as a saprophyte or a parasite.
Figures

References
-
- Barondes S H, Cooper D N, Gitt M A, Leffler H. Galectins. Structure and function of a large family of animal lectins. J Biol Chem. 1994;269:20807–20810. - PubMed
-
- Cassone A, Torosantucci A, Boccanera M, Pelligrini G, Palma C, Malavasi F. Production and characterization of a monoclonal antibody to a cell-surface, glucomannoprotein constituent of Candida albicans and other pathogenic Candida species. J Med Microbiol. 1988;27:233–238. - PubMed
-
- Czop J K, Austen K F. A beta-glucan inhibitable receptor on human monocytes: its identity with the phagocytic receptor for particulate activators of the alternative complement pathway. J Immunol. 1985;134:2588–2593. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
