

Natural interferon alpha/beta-producing cells link innate and adaptive immunity
- PMID: 10899908
- PMCID: PMC2193254
- DOI: 10.1084/jem.192.2.219
Natural interferon alpha/beta-producing cells link innate and adaptive immunity
Abstract
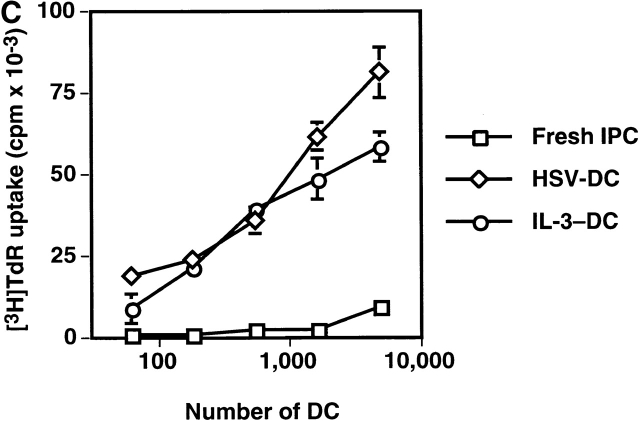
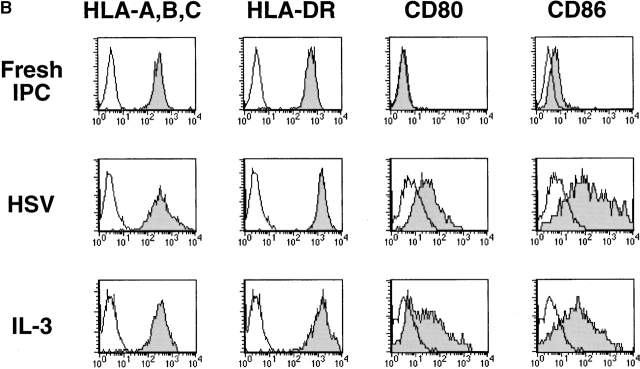
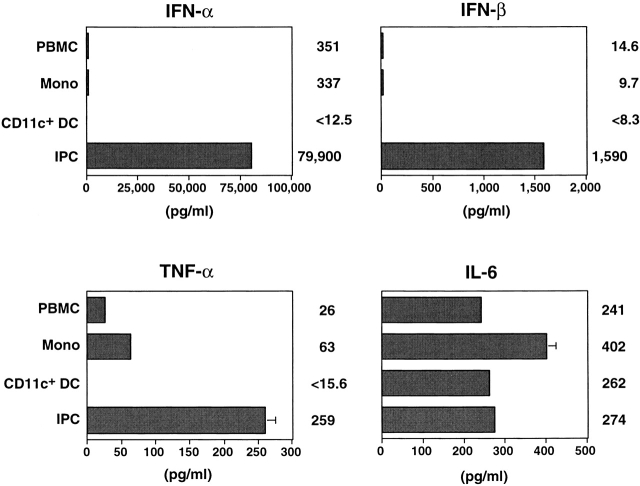
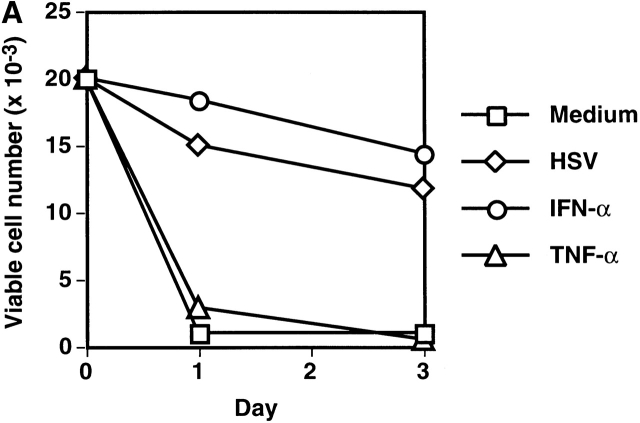
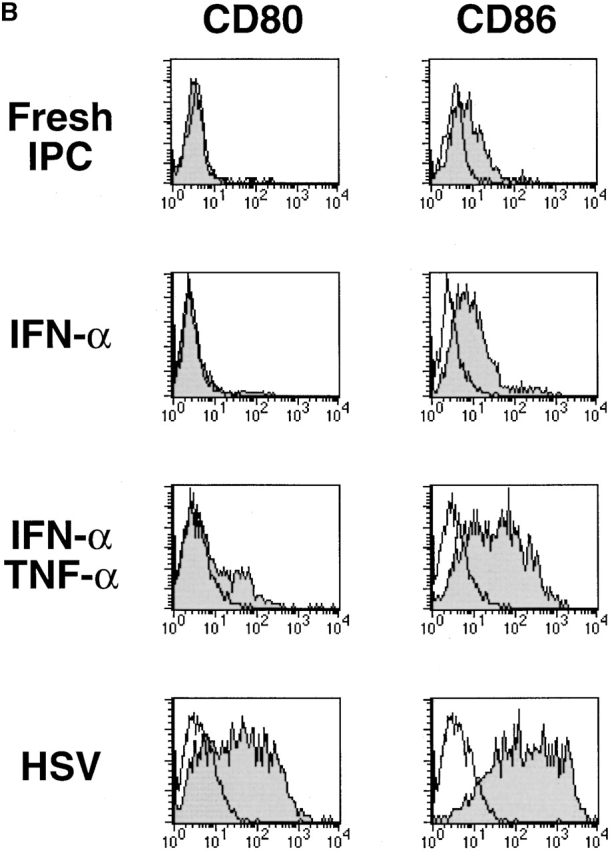
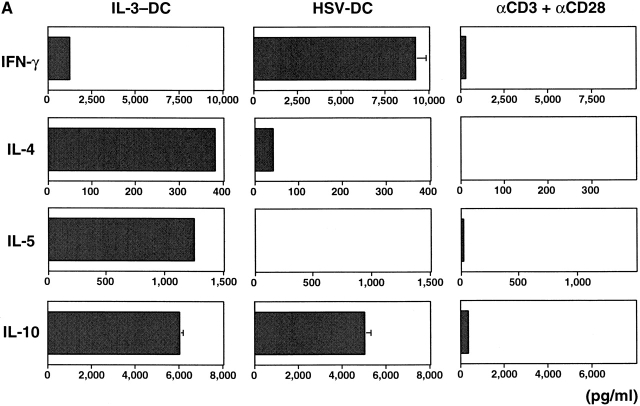
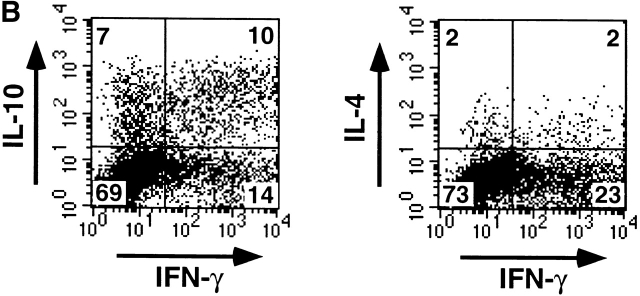
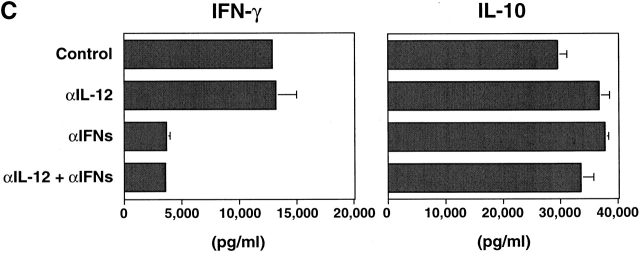
Innate immune responses to pathogens critically impact the development of adaptive immune responses. However, it is not completely understood how innate immunity controls the initiation of adaptive immunities or how it determines which type of adaptive immunity will be induced to eliminate a given pathogen. Here we show that viral stimulation not only triggers natural interferon (IFN)-alpha/beta-producing cells (IPCs) to produce vast amounts of antiviral IFN-alpha/beta but also induces these cells to differentiate into dendritic cells (DCs). IFN-alpha/beta and tumor necrosis factor alpha produced by virus-activated IPCs act as autocrine survival and DC differentiation factors, respectively. The virus-induced DCs stimulate naive CD4(+) T cells to produce IFN-gamma and interleukin (IL)-10, in contrast to IL-3-induced DCs, which stimulate naive CD4(+) T cells to produce T helper type 2 cytokines IL-4, IL-5, and IL-10. Thus, IPCs may play two master roles in antiviral immune responses: directly inhibiting viral replication by producing large amounts of IFN-alpha/beta, and subsequently triggering adaptive T cell-mediated immunity by differentiating into DCs. IPCs constitute a critical link between innate and adaptive immunity.
Figures

References
-
- Fearon D.T., Locksley R.M. The instructive role of innate immunity in the acquired immune response. Science. 1996;272:50–53. - PubMed
-
- Medzhitov R., Janeway C.A., Jr. Innate immunityimpact on the adaptive immune response. Curr. Opin. Immunol. 1997;9:4–9. - PubMed
-
- Sallusto F., Cella M., Danieli C., Lanzavecchia A. Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartmentdownregulation by cytokines and bacterial products. J. Exp. Med. 1995;182:389–400. - PMC - PubMed
-
- Cella M., Engering A., Pinet V., Pieters J., Lanzavecchia A. Inflammatory stimuli induce accumulation of MHC class II complexes on dendritic cells. Nature. 1997;388:782–787. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
