

Using chimeric hypoviruses to fine-tune the interaction between a pathogenic fungus and its plant host
- PMID: 10906210
- PMCID: PMC112277
- DOI: 10.1128/jvi.74.16.7562-7567.2000
Using chimeric hypoviruses to fine-tune the interaction between a pathogenic fungus and its plant host
Abstract
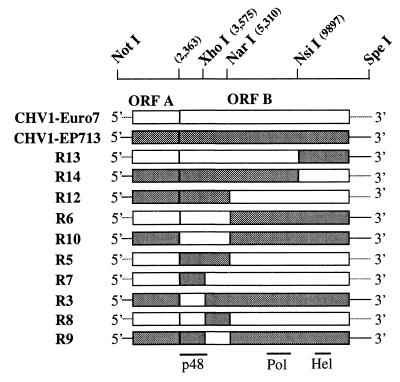
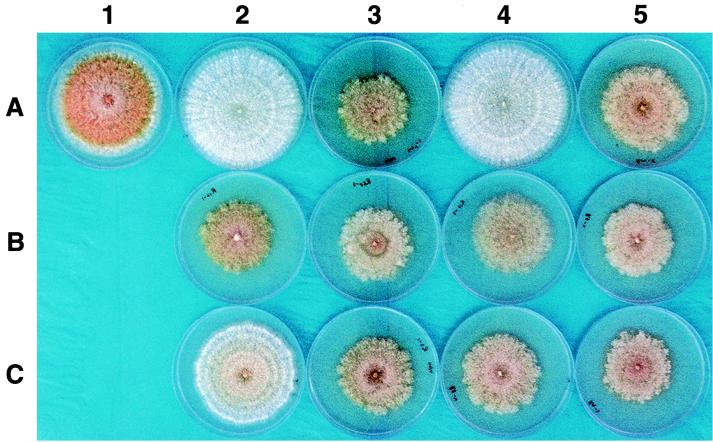
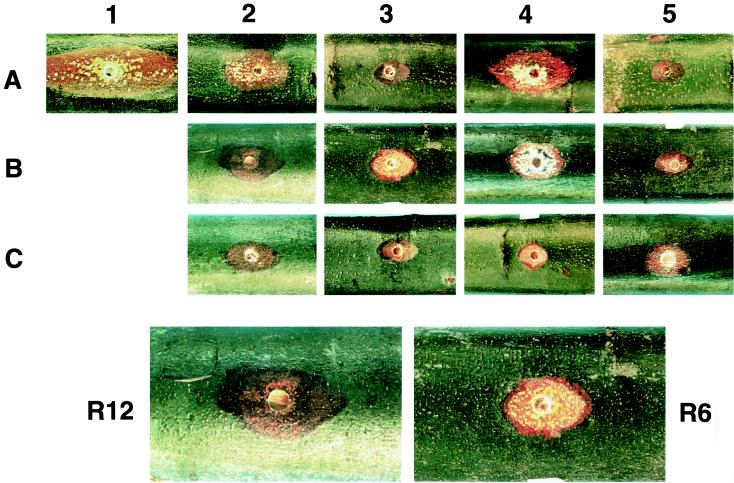
Infectious cDNA clones of mild (CHV1-Euro7) and severe (CHV1-EP713) hypovirus strains responsible for virulence attenuation (hypovirulence) of the chestnut blight fungus Cryphonectria parasitica were used to construct viable chimeric viruses. Differences in virus-mediated alterations of fungal colony morphology, growth rate, and canker morphology were mapped to a region of open reading frame B extending from nucleotides 2,363 to 9, 904. By swapping domains within this region, it was possible to generate chimeric hypovirus-infected C. parasitica isolates that exhibited a spectrum of defined colony and canker morphologies. Several severe strain traits were observed to be dominant. It was also possible to uncouple the severe strain traits of small canker size and suppression of asexual sporulation. For example, fungal isolates infected with a chimera containing nucleotides 2363 through 5310 from CHV1-Euro7 in a CHV1-713 background formed small cankers that were similar in size to that caused by CHV1-EP713-infected isolates but with the capacity for producing asexual spores at levels approaching that observed for fungal isolates infected with the mild strain. These results demonstrate that hypoviruses can be engineered to fine-tune the interaction between a pathogenic fungus and its plant host. The identification of specific hypovirus domains that differentially contribute to canker morphology and sporulation levels also provides considerable utility for continuing efforts to enhance biological control potential by balancing hypovirulence and ecological fitness.
Figures
References
-
- Anagnostakis S L. Biological control of chestnut blight. Science. 1982;215:466–471. - PubMed
-
- Anagnostakis S L, Day P R. Hypovirulence conversion in Endothia parasitica. Phytopathology. 1979;69:1226–1229.
-
- Bissegger M, Rigling D, Heiniger U. Population structure and disease development of Cryphonectria parasitica in European chestnut forests in the presence of natural hypovirulence. Phytopathology. 1997;87:50–59. - PubMed
-
- Brandy B P, Tavantzis S M. Effect of hypovirulent Rhizoctonia solani on rhizoctonia disease, growth, and development. Am Potato J. 1990;67:189–199.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
