

Similarities and differences in the conformation of protein-DNA complexes at the U1 and U6 snRNA gene promoters
- PMID: 10908334
- PMCID: PMC102643
- DOI: 10.1093/nar/28.14.2771
Similarities and differences in the conformation of protein-DNA complexes at the U1 and U6 snRNA gene promoters
Abstract
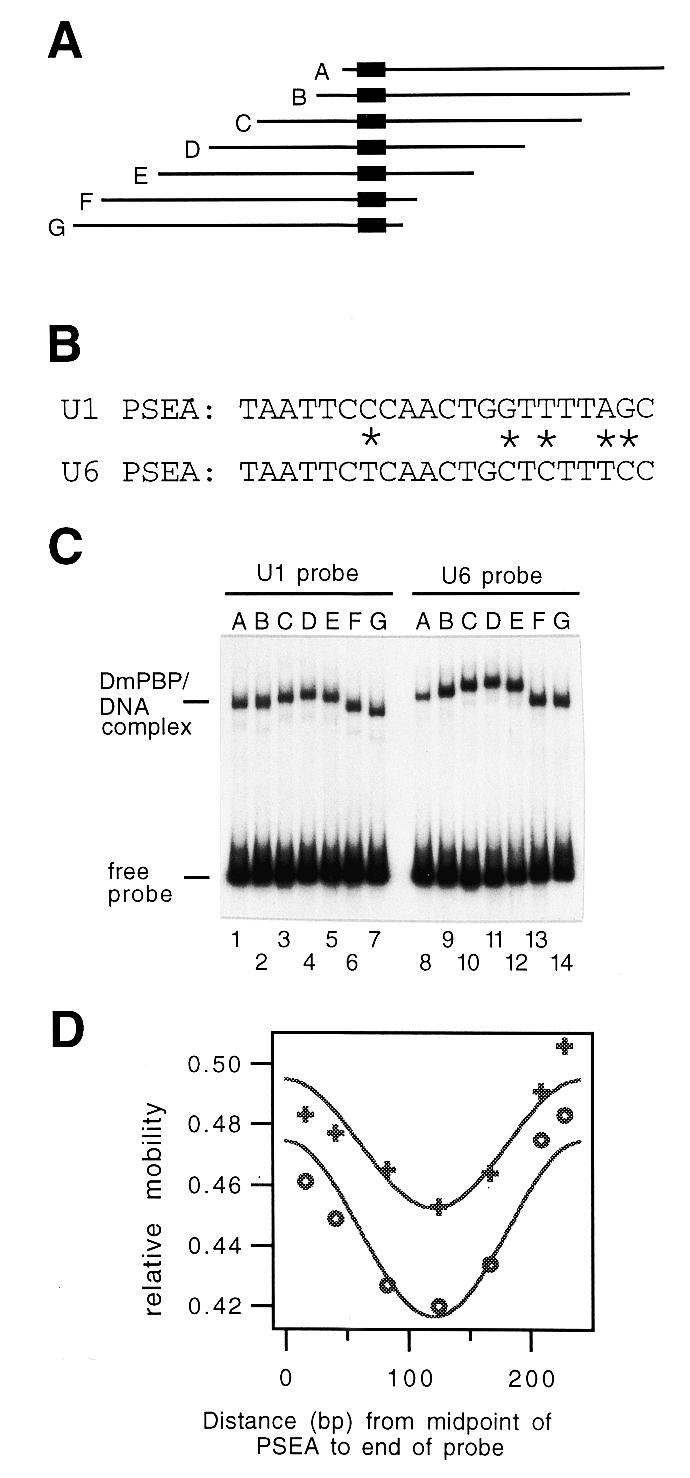
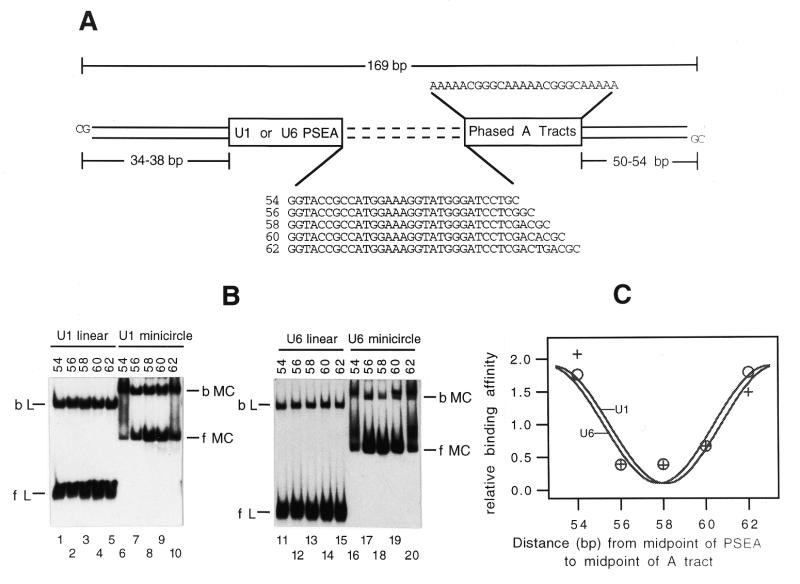
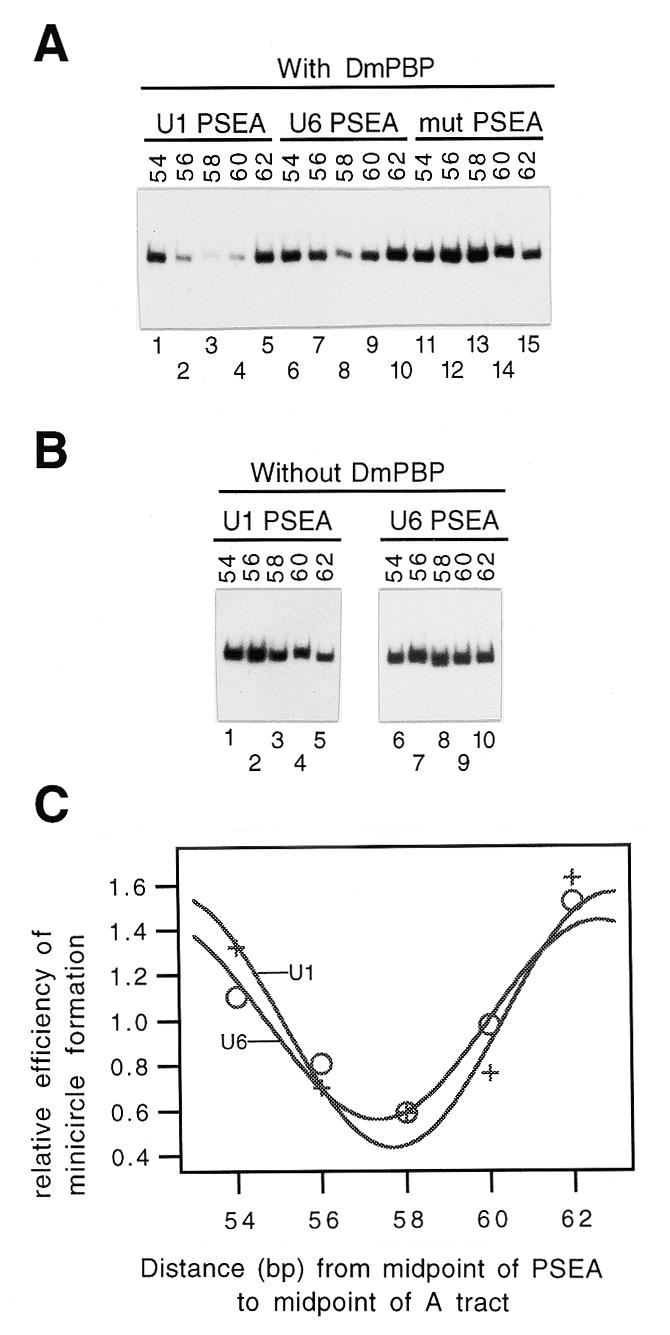
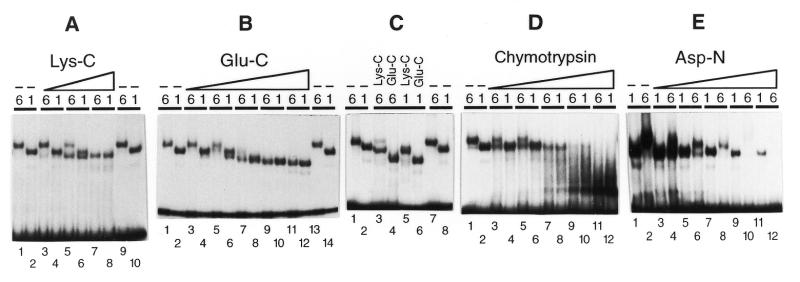
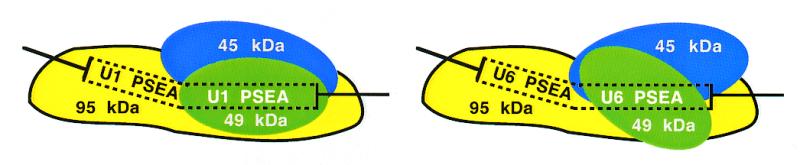
Most small nuclear RNAs (snRNAs) are synthesized by RNA polymerase II, but U6 snRNA is synthesized by RNA polymerase III. In the fruit fly Drosophila melanogaster the RNA polymerase specificity of the snRNA genes is determined by a few nucleotide differences within the proximal sequence element (PSE), a conserved sequence located approximately 40-65 bp upstream of the transcription start site. The PSE is essential for transcription of both RNA polymerase II-transcribed and RNA polymerase III-transcribed snRNA genes and is recognized in Drosophila by a multi-subunit protein factor termed DM:PBP. Previous studies that employed site-specific protein-DNA photocrosslinking indicated that the conformation of the DNA-protein complex is different depending upon whether DM:PBP is bound to a U1 or U6 PSE sequence. These conformational differences of the complex probably represent an early step in determining the selection of the correct RNA polymerase. We have now obtained evidence that DM:PBP modestly bends the DNA upon interacting with the PSE and that the direction of DNA bending is similar for both the U1 and U6 PSEs. Under the assumption that DM:PBP does not significantly twist the DNA, the direction of the bend in both cases is toward the face of the DNA helix contacted by the 45 kDa subunit of DM:PBP. Together with data from partial proteolysis assays, these results indicate that the conformational differences in the complexes of DM:PBP with the U1 and U6 PSEs more likely occur at the protein level rather than at the DNA level.
Figures
References
-
- Hernandez N. (1992) In McKnight,S.L. and Yamamoto,K.R. (eds), Transcriptional Regulation. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 281–313.
-
- Lobo S.M. and Hernandez,N.T. (1994) In Conaway,R.C. and Conaway,J.W. (eds), Transcription: Mechanisms and Regulation. Raven Press, New York, NY, pp. 127–159.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
