

The SNARE Vti1a-beta is localized to small synaptic vesicles and participates in a novel SNARE complex
- PMID: 10908612
- PMCID: PMC6772562
- DOI: 10.1523/JNEUROSCI.20-15-05724.2000
The SNARE Vti1a-beta is localized to small synaptic vesicles and participates in a novel SNARE complex
Abstract
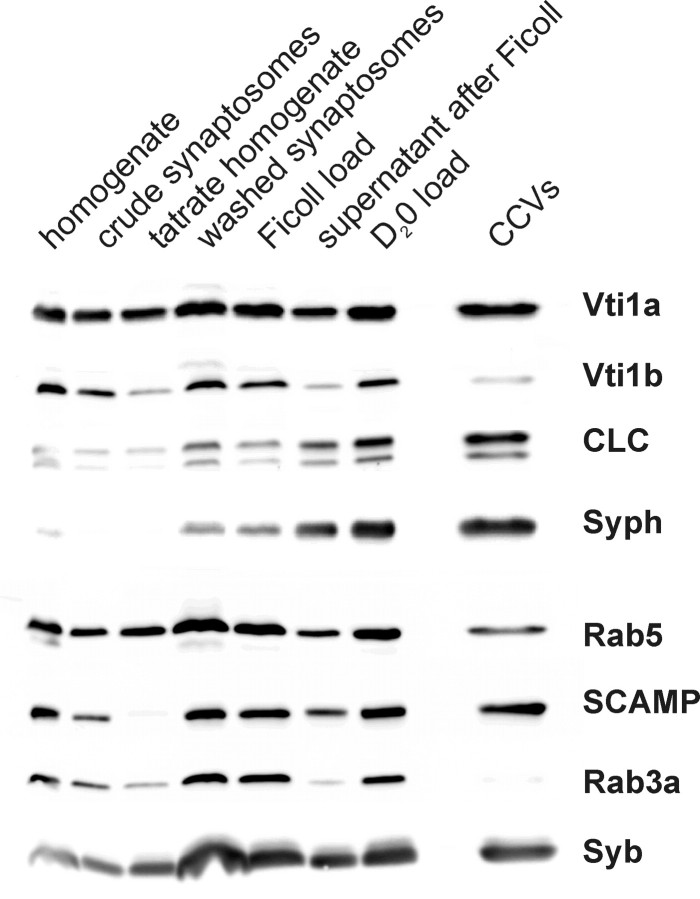
Specific soluble N-ethylmaleimide-sensitive factor attachment protein (SNAP) receptor (SNARE) proteins are required for different membrane transport steps. The SNARE Vti1a has been colocalized with Golgi markers and Vti1b with Golgi and the trans-Golgi network or endosomal markers in fibroblast cell lines. Here we study the distribution of Vti1a and Vti1b in brain. Vti1b was found in synaptic vesicles but was not enriched in this organelle. A brain-specific splice variant of Vti1a was identified that had an insertion of seven amino acid residues next to the putative SNARE-interacting helix. This Vti1a-beta was enriched in small synaptic vesicles and clathrin-coated vesicles isolated from nerve terminals. Vti1a-beta also copurified with the synaptic vesicle R-SNARE synaptobrevin during immunoisolation of synaptic vesicles and endosomes. Therefore, both synaptobrevin and Vti1a-beta are integral parts of synaptic vesicles throughout their life cycle. Vti1a-beta was part of a SNARE complex in nerve terminals, which bound N-ethylmaleimide-sensitive factor and alpha-SNAP. This SNARE complex was different from the exocytic SNARE complex because Vti1a-beta was not coimmunoprecipitated with syntaxin 1 or SNAP-25. These data suggest that Vti1a-beta does not function in exocytosis but in a separate SNARE complex in a membrane fusion step during recycling or biogenesis of synaptic vesicles.
Figures










References
-
- Advani RJ, Bae HR, Bock JB, Chao DS, Doung YC, Prekeris R, Yoo JS, Scheller RH. Seven novel mammalian SNARE proteins localize to distinct membrane compartments. J Biol Chem. 1998;273:10317–10324. - PubMed
-
- Alés E, Tabares L, Poyato JM, Valero V, Lindau M, de Toledo GA. High calcium concentrations shift the mode of exocytosis to the kiss-and-run mechanism. Nat Cell Biol. 1999;1:40–44. - PubMed
-
- Barnstable CJ, Hofstein R, Akagawa K. A marker of early amacrine cell development in rat retina. Dev Brain Res. 1985;20:286–290. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases