

Depletion of a microtubule-associated motor protein induces the loss of dendritic identity
- PMID: 10908619
- PMCID: PMC6772545
- DOI: 10.1523/JNEUROSCI.20-15-05782.2000
Depletion of a microtubule-associated motor protein induces the loss of dendritic identity
Abstract
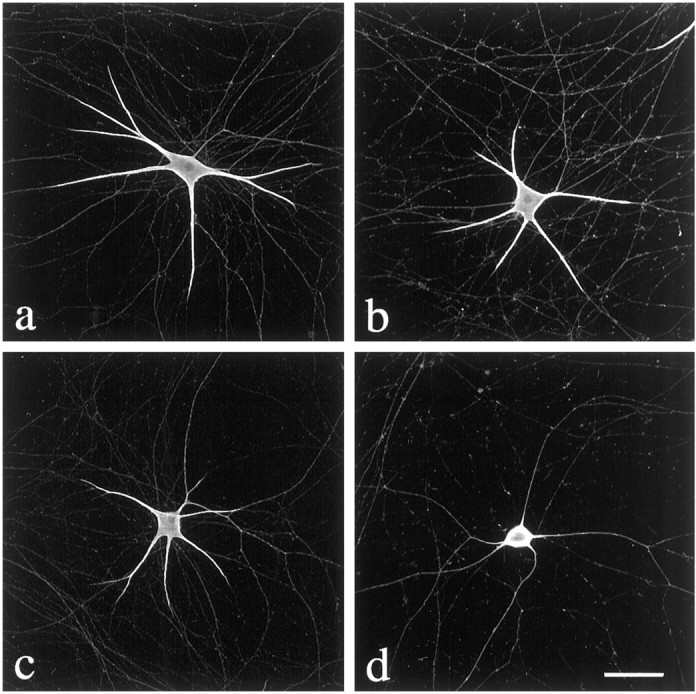
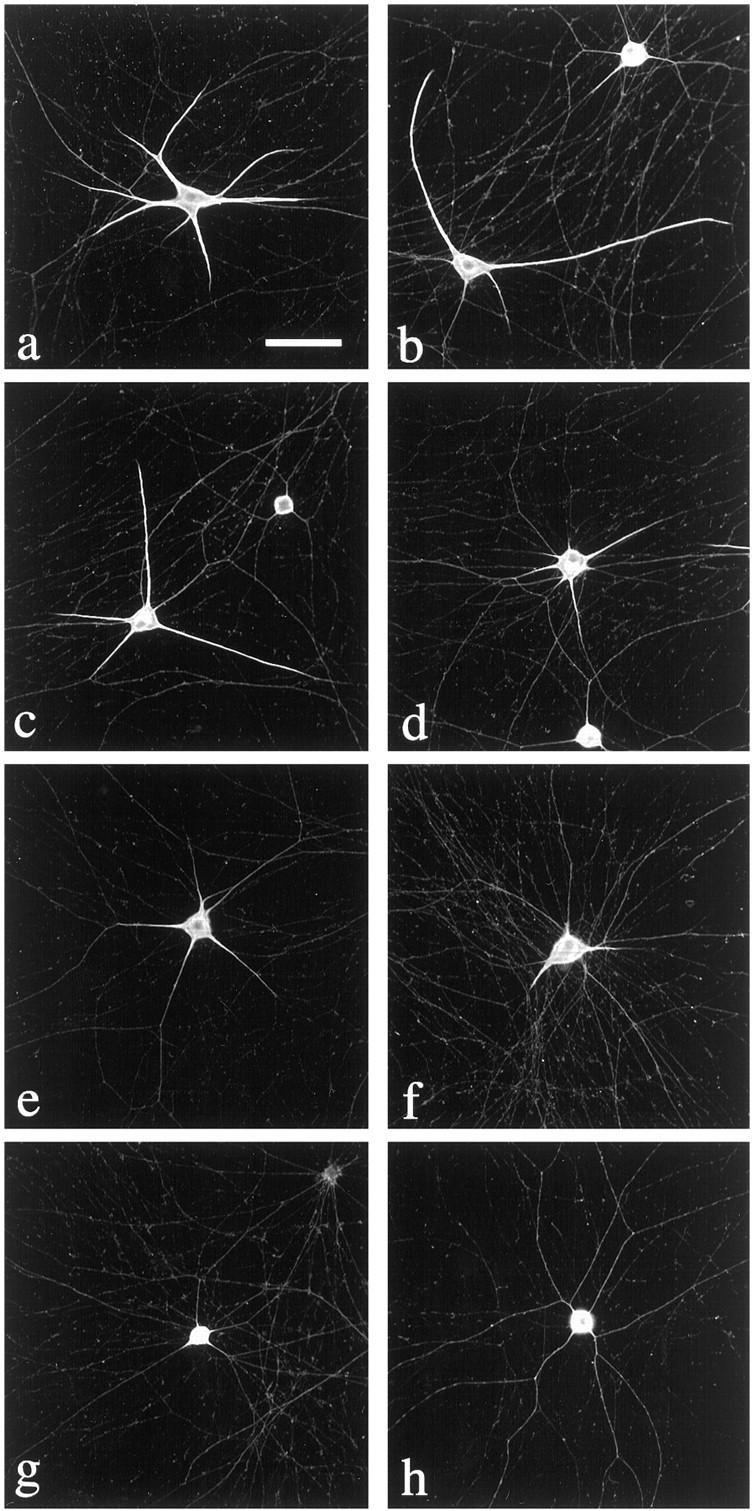
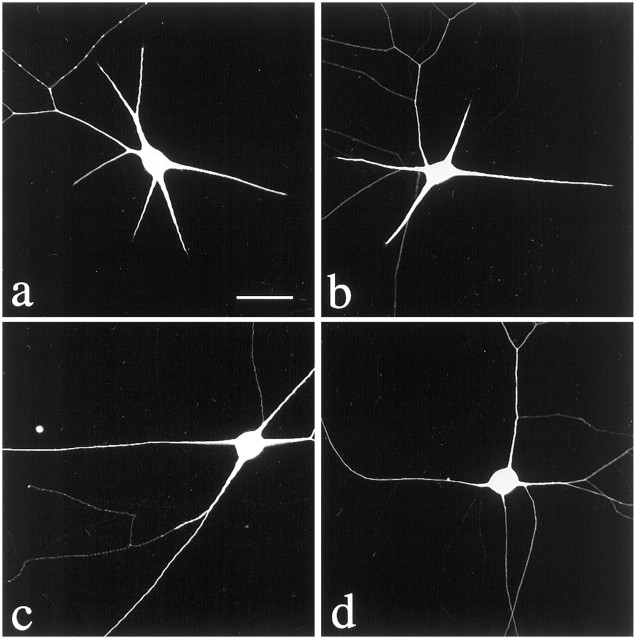
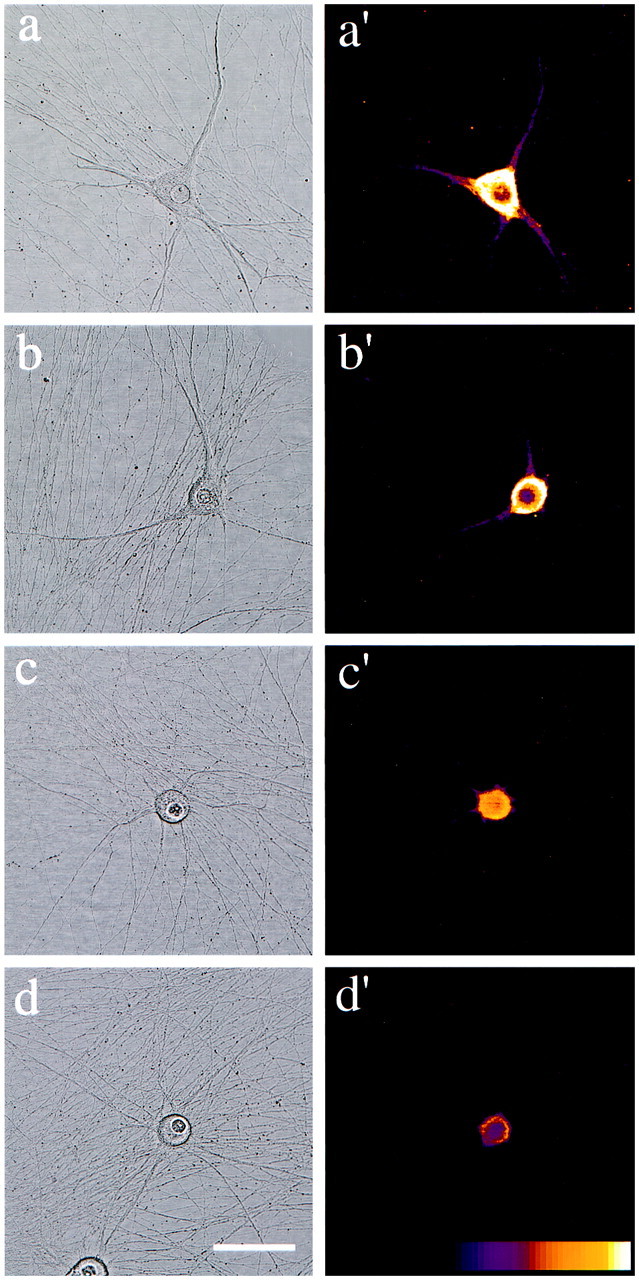
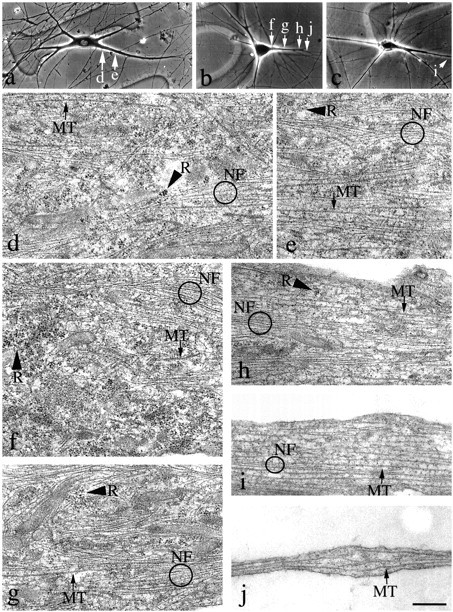
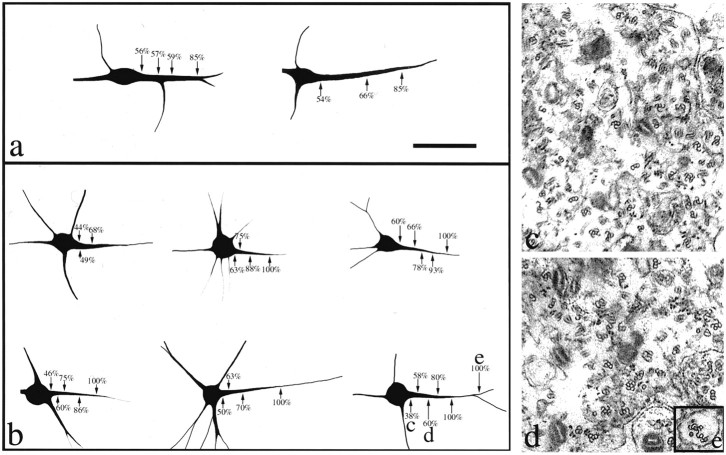
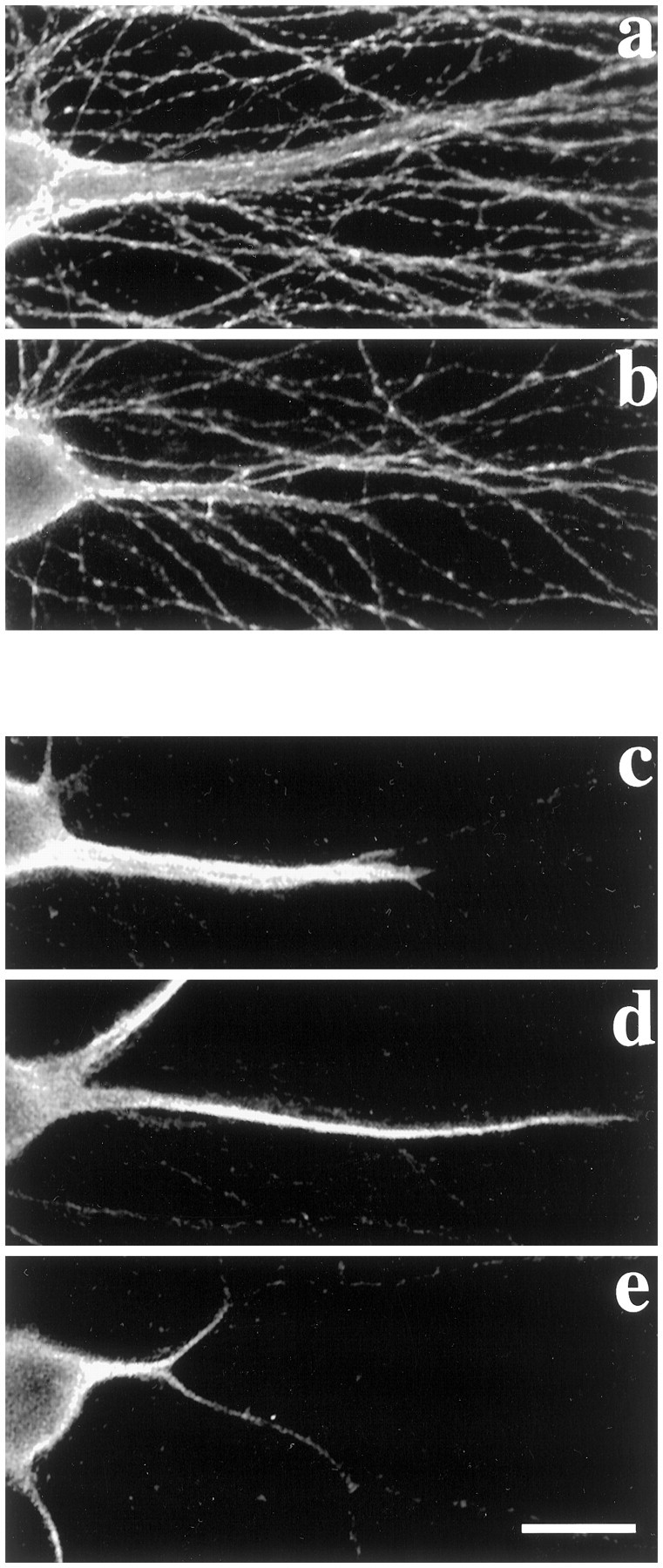
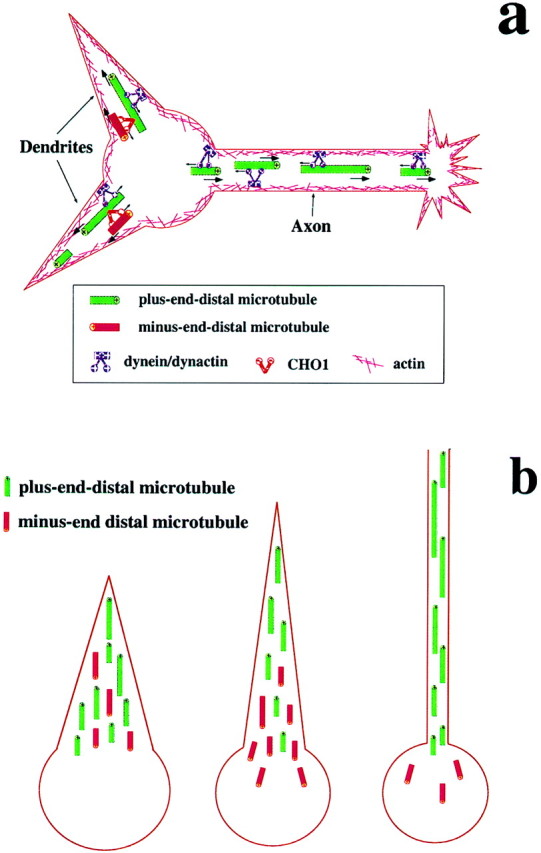
Dendrites are short stout tapering processes that are rich in ribosomes and Golgi elements, whereas axons are long thin processes of uniform diameter that are deficient in these organelles. It has been hypothesized that the unique morphological and compositional features of axons and dendrites result from their distinct patterns of microtubule polarity orientation. The microtubules within axons are uniformly oriented with their plus ends distal to the cell body, whereas microtubules within dendrites are nonuniformly oriented. The minus-end-distal microtubules are thought to arise via their specific transport into dendrites by the motor protein known as CHO1/MKLP1. According to this model, CHO1/MKLP1 transports microtubules with their minus ends leading into dendrites by generating forces against the plus-end-distal microtubules, thus creating drag on the plus-end-distal microtubules. Here we show that depletion of CHO1/MKLP1 from cultured neurons causes a rapid redistribution of microtubules within dendrites such that minus-end-distal microtubules are chased back to the cell body while plus-end-distal microtubules are redistributed forward. The dendrite grows significantly longer and thinner, loses its taper, and acquires a progressively more axon-like organelle composition. These results suggest that the forces generated by CHO1/MKLP1 are necessary for maintaining the minus-end-distal microtubules in the dendrite, for antagonizing the anterograde transport of the plus-end-distal microtubules, and for sustaining a pattern of microtubule organization necessary for the maintenance of dendritic morphology and composition. Thus, we would conclude that dendritic identity is dependent on forces generated by CHO1/MKLP1.
Figures
References
-
- Ahmad FJ, Hughey J, Wittmann T, Hyman A, Greaser M, Baas PW. Motor proteins regulate force interactions between microtubules and microfilaments in the axon. Nat Cell Biol. 2000;2:276–280. - PubMed
-
- Baas PW. Microtubules and neuronal polarity: lessons from mitosis. Neuron. 1999;22:23–31. - PubMed
-
- Baas PW. Strategies for studying microtubule transport in the neuron. Microsc Res Tech. 2000;48:75–84. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources