

Molecular basis of protein kinase C-induced activation of ATP-sensitive potassium channels
- PMID: 10908656
- PMCID: PMC16821
- DOI: 10.1073/pnas.160068997
Molecular basis of protein kinase C-induced activation of ATP-sensitive potassium channels
Abstract
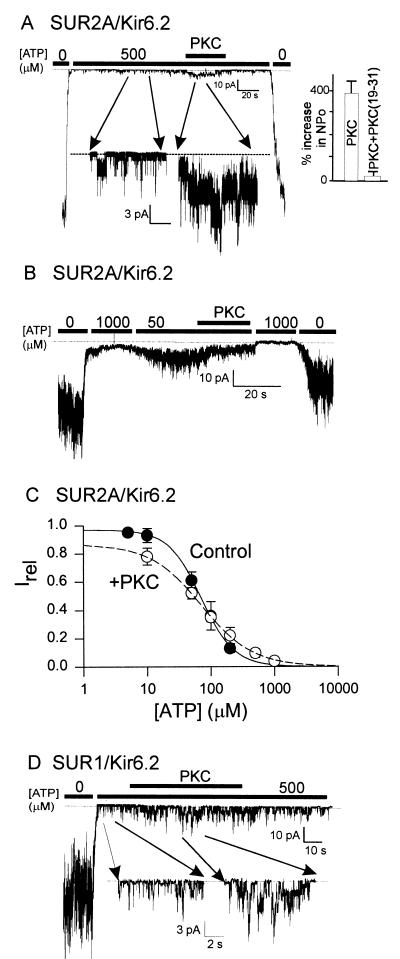
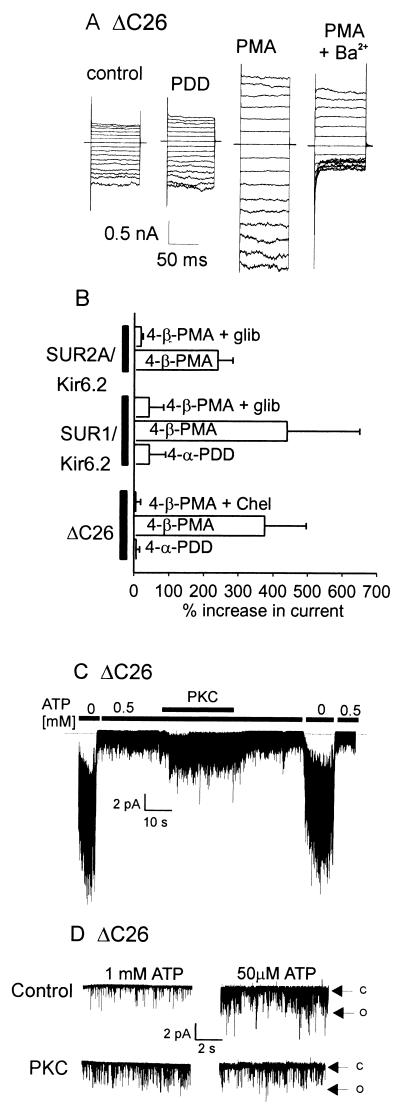
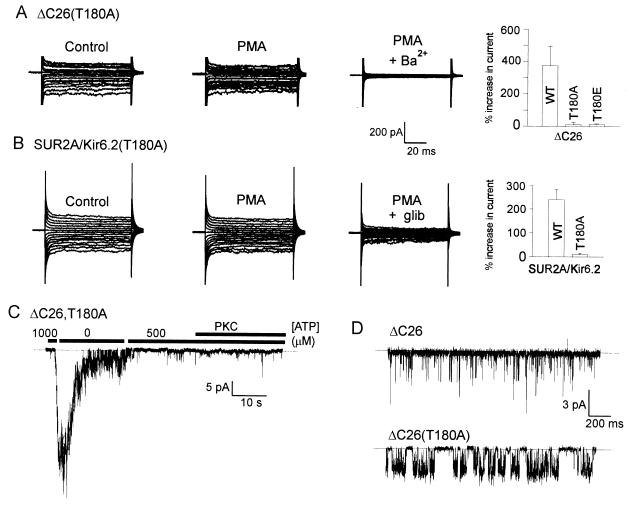
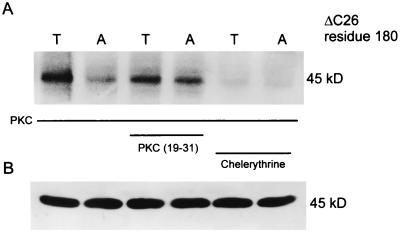
Potassium channels that are inhibited by internal ATP (K(ATP) channels) provide a critical link between metabolism and cellular excitability. Protein kinase C (PKC) acts on K(ATP) channels to regulate diverse cellular processes, including cardioprotection by ischemic preconditioning and pancreatic insulin secretion. PKC action decreases the Hill coefficient of ATP binding to cardiac K(ATP) channels, thereby increasing their open probability at physiological ATP concentrations. We show that PKC similarly regulates recombinant channels from both the pancreas and heart. Surprisingly, PKC acts via phosphorylation of a specific, conserved threonine residue (T180) in the pore-forming subunit (Kir6.2). Additional PKC consensus sites exist on both Kir and the larger sulfonylurea receptor (SUR) subunits. Nonetheless, T180 controls changes in open probability induced by direct PKC action either in the absence of, or in complex with, the accessory SUR1 (pancreatic) or SUR2A (cardiac) subunits. The high degree of conservation of this site among different K(ATP) channel isoforms suggests that this pathway may have wide significance for the physiological regulation of K(ATP) channels in various tissues and organelles.
Figures

References
-
- Seino S. Annu Rev Physiol. 1999;61:337–362. - PubMed
-
- Babenko A P, Aguilar-Bryan L, Bryan J. Annu Rev Physiol. 1998;60:667–687. - PubMed
-
- Aguilar-Bryan L, Bryan J. Endocr Rev. 1999;20:101–135. - PubMed
-
- Inagaki N, Gonoi T, Clement J P, IV, Namba N, Inazawa J, Gonzalez G, Aguilar Bryan L, Seino S, Bryan J. Science. 1995;270:1166–1170. - PubMed
-
- Inagaki N, Gonoi T, Clement J P, IV, Wang C-Z, Aguilar-Bryan L, Bryan J, Seino S. Neuron. 1996;16:1011–1017. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
