

Specific binding sites for alcohols and anesthetics on ligand-gated ion channels
- PMID: 10908659
- PMCID: PMC16863
- DOI: 10.1073/pnas.160128797
Specific binding sites for alcohols and anesthetics on ligand-gated ion channels
Abstract
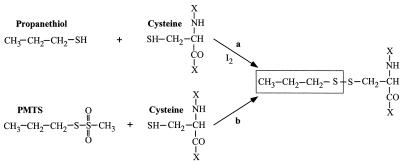
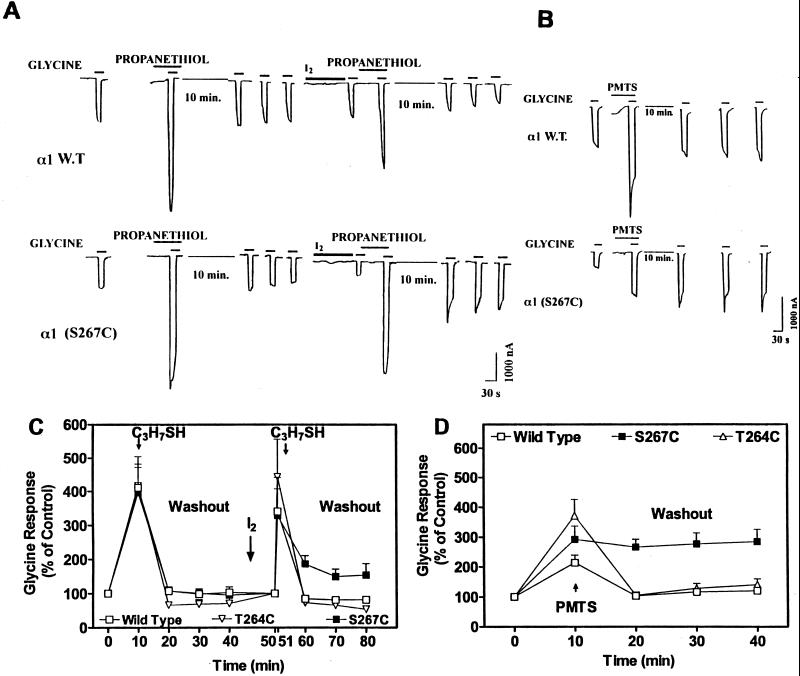
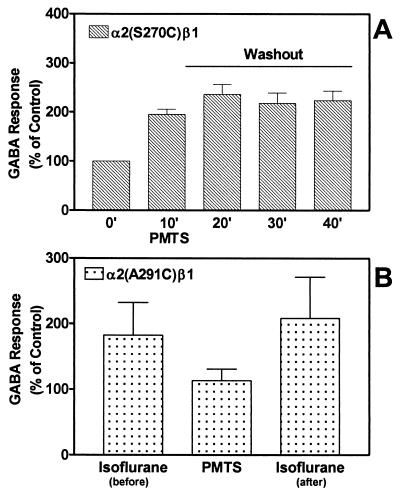
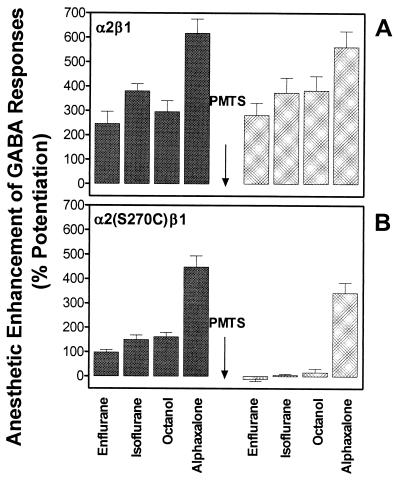
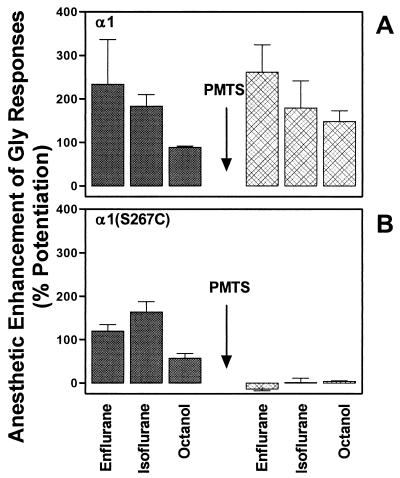
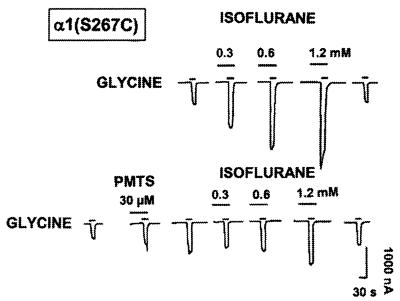
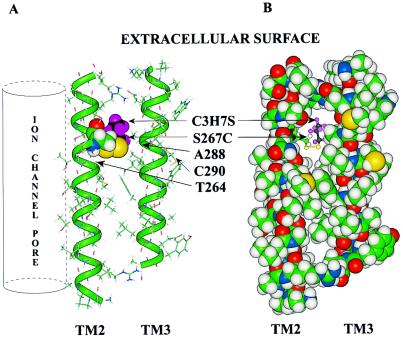
Ligand-gated ion channels are a target for inhaled anesthetics and alcohols in the central nervous system. The inhibitory strychnine-sensitive glycine and gamma-aminobutyric acid type A receptors are positively modulated by anesthetics and alcohols, and site-directed mutagenesis techniques have identified amino acid residues important for the action of volatile anesthetics and alcohols in these receptors. A key question is whether these amino acids are part of an alcohol/anesthetic-binding site. In the present study, we used an alkanethiol anesthetic to covalently label its binding site by mutating selected amino acids to cysteine. We demonstrated that the anesthetic propanethiol, or alternatively, propyl methanethiosulfonate, covalently binds to cysteine residues introduced into a specific second transmembrane site in glycine receptor and gamma-aminobutyric acid type A receptor subunits and irreversibly enhances receptor function. Moreover, upon permanent occupation of the site by propyl disulfide, the usual ability of octanol, enflurane, and isoflurane to potentiate the function of the ion channels was lost. This approach provides strong evidence that the actions of anesthetics in these receptors are due to binding at a single site.
Figures
References
-
- Kennedy S K, Longnecker D E. In: The Pharmacological Basis of Therapeutics. Moliniff P B, Ruddon R W, editors. New York: McGraw–Hill; 1996. pp. 304–305.
-
- Meyer H H. Arch Exp Pathol Pharmakol. 1899;42:109–118.
-
- Overton E. Studien uber die Narkose, Zugleich ein Beitrag zur Allgemeinen Pharmakologie. Jena, Germany: Gustav Fischer; 1901. pp. 1–195.
-
- Cantor R S. Biochemistry. 1997;36:2339–2344. - PubMed
-
- Forman S A, Miller K W, Yellen G. Mol Pharmacol. 1995;48:574–581. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
