

Characterization of LrpC DNA-binding properties and regulation of Bacillus subtilis lrpC gene expression
- PMID: 10913073
- PMCID: PMC94611
- DOI: 10.1128/JB.182.16.4414-4424.2000
Characterization of LrpC DNA-binding properties and regulation of Bacillus subtilis lrpC gene expression
Abstract
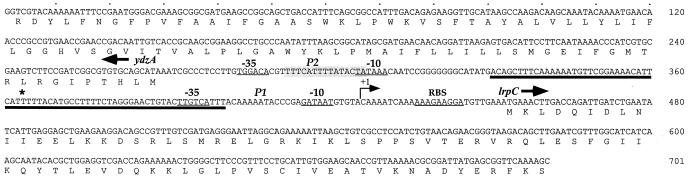
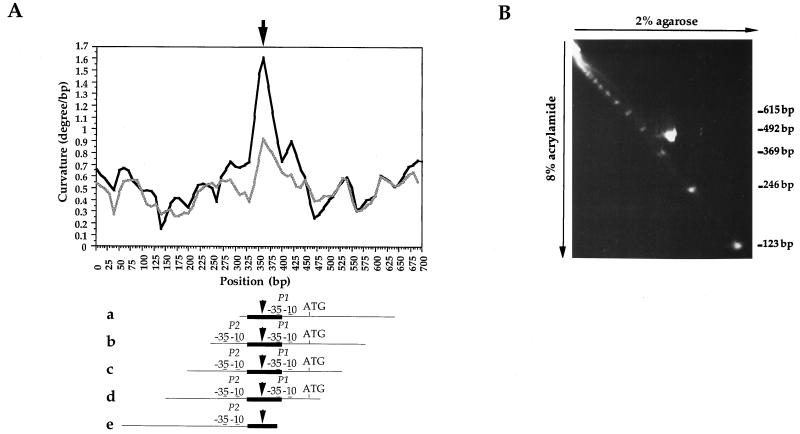
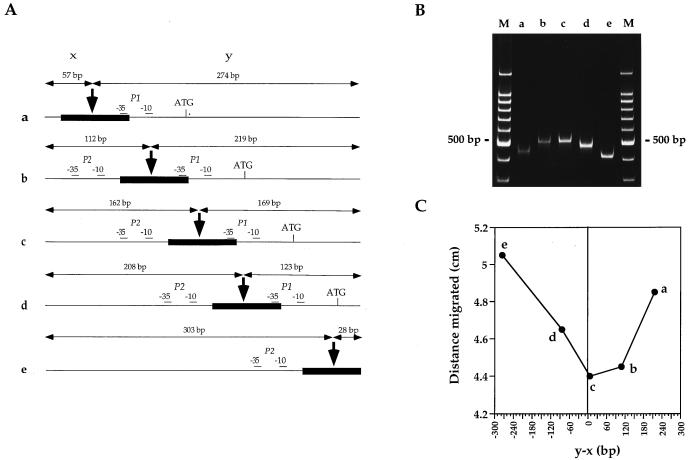
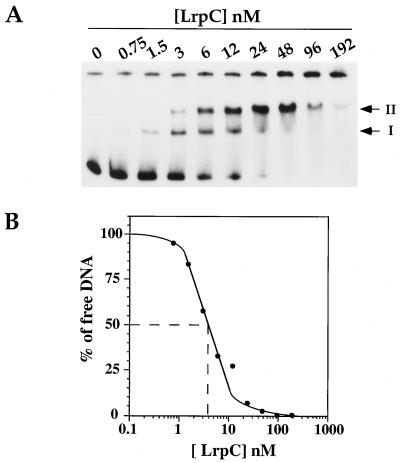
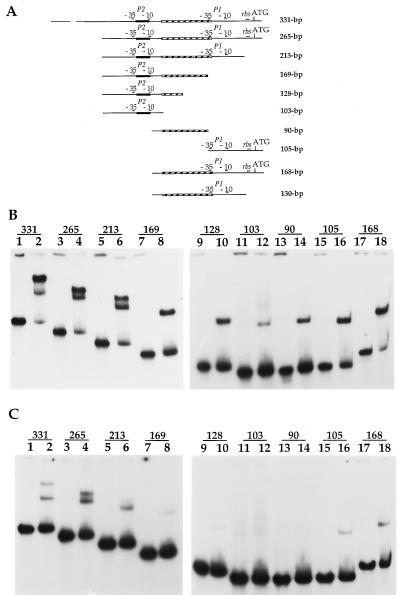
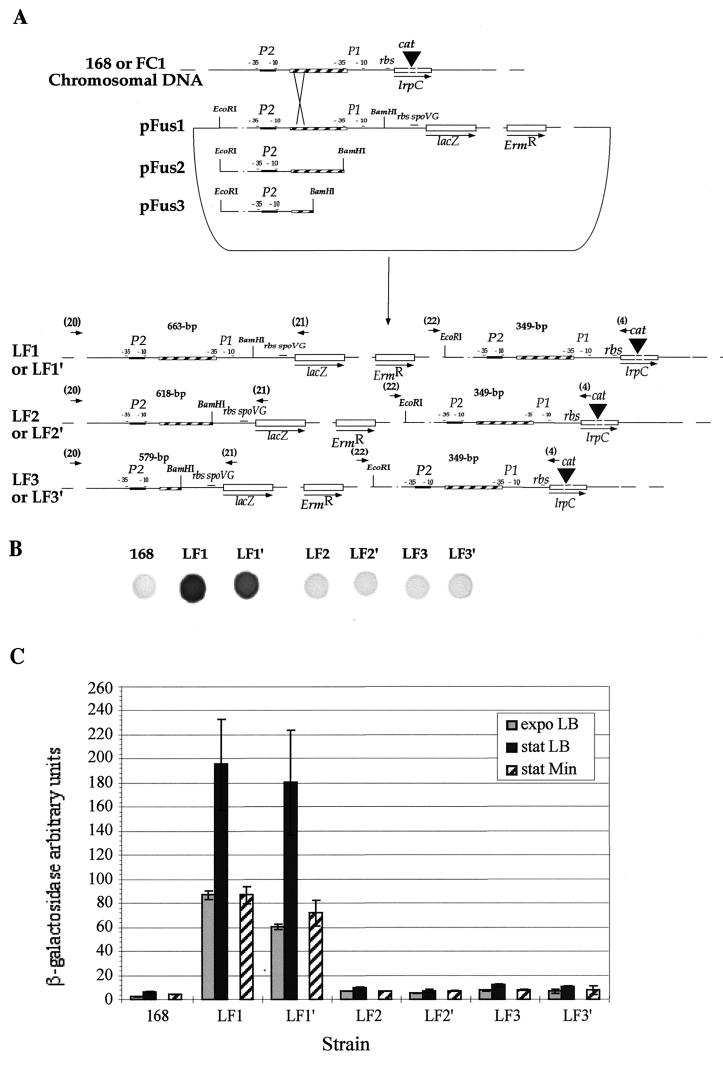
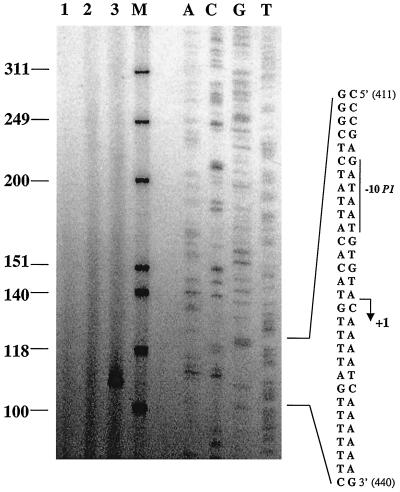
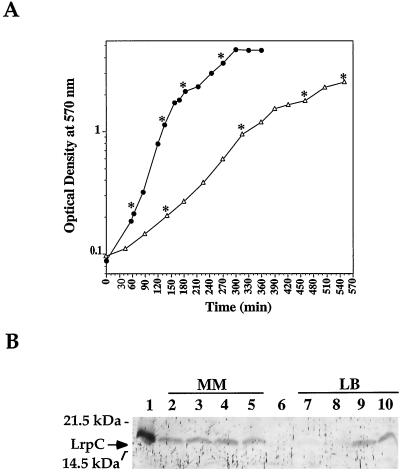
The lrpC gene was identified during the Bacillus subtilis genome sequencing project. Previous experiments suggested that LrpC has a role in sporulation and in the regulation of amino acid metabolism and that it shares features with Escherichia coli Lrp, a transcription regulator (C. Beloin, S. Ayora, R. Exley, L. Hirschbein, N. Ogasawara, Y. Kasahara, J. C. Alonso, and F. Le Hégarat, Mol. Gen. Genet. 256:63-71, 1997). To characterize the interactions of LrpC with DNA, the protein was overproduced and purified. We show that LrpC binds to multiple sites in the upstream region of its own gene with a stronger affinity for a region encompassing P1, one of the putative promoters identified (P1 and P2). By analyzing lrpC-lacZ transcriptional fusions, we demonstrated that P1 is the major in vivo promoter and that, unlike many members of the lrp/asnC family, lrpC is not negatively autoregulated but rather slightly positively autoregulated. Production of LrpC in vivo is low in both rich and minimal media (50 to 300 LrpC molecules per cell). In rich medium, the cellular LrpC content is six- to sevenfold lower during the exponentional phase than during the stationary growth phase. Possible determinants and the biological significance of the regulation of lrpC expression are discussed.
Figures
References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Beloin C, Ayora S, Exley R, Hirschbein L, Ogasawara N, Kasahara Y, Alonso J C, Le Hégarat F. Characterization of an lrp-like (lrpC) gene from Bacillus subtilis. Mol Gen Genet. 1997;256:63–71. - PubMed
-
- Brennan R G, Matthews B W. The helix-turn-helix DNA binding motif. J Biol Chem. 1989;264:1903–1906. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
