

A Cl(-) cotransporter selective for NH(4)(+) over K(+) in glial cells of bee retina
- PMID: 10919861
- PMCID: PMC2229498
- DOI: 10.1085/jgp.116.2.125
A Cl(-) cotransporter selective for NH(4)(+) over K(+) in glial cells of bee retina
Abstract
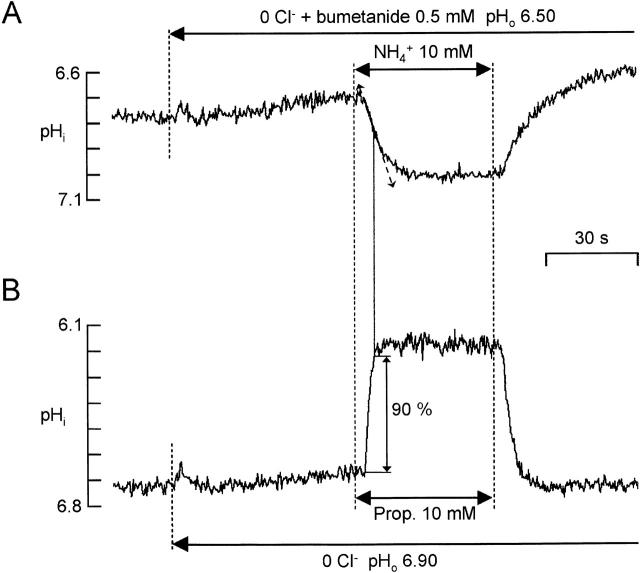
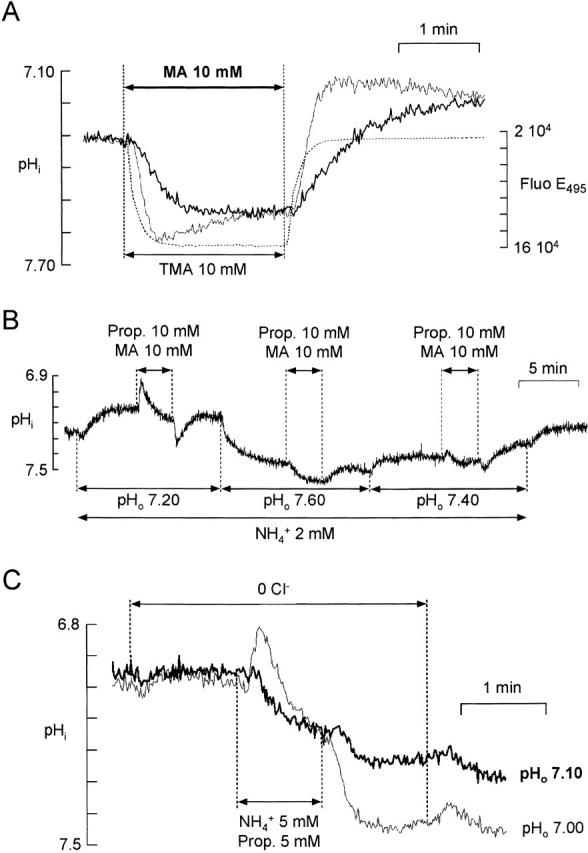
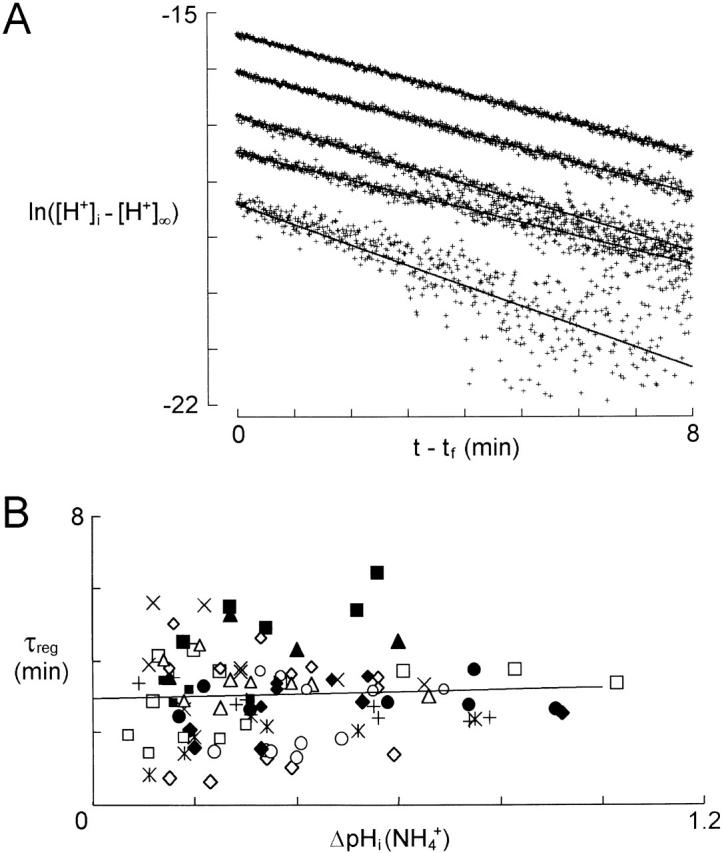
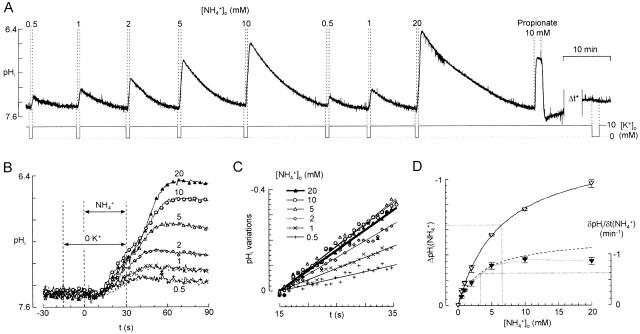
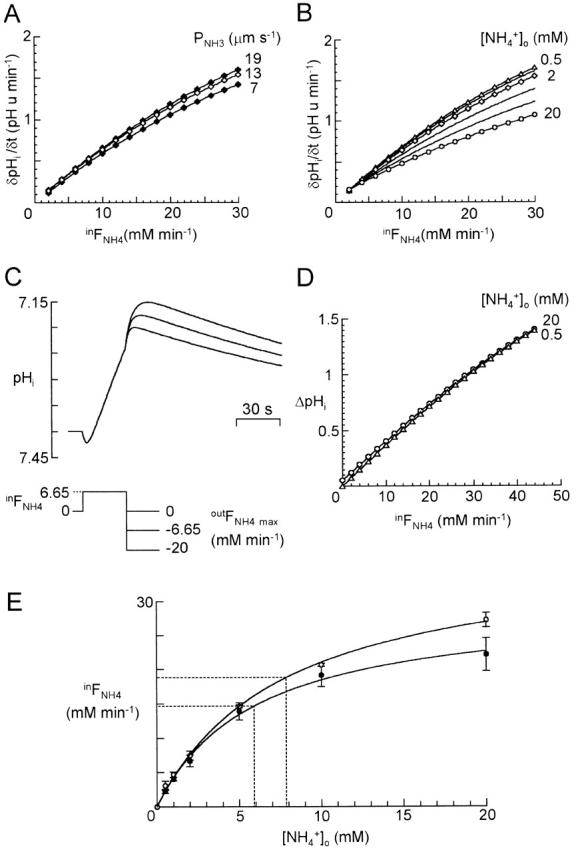
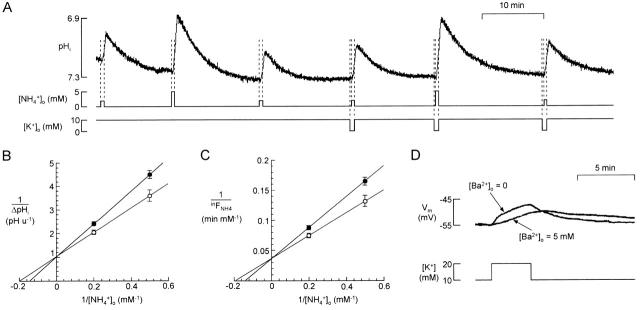
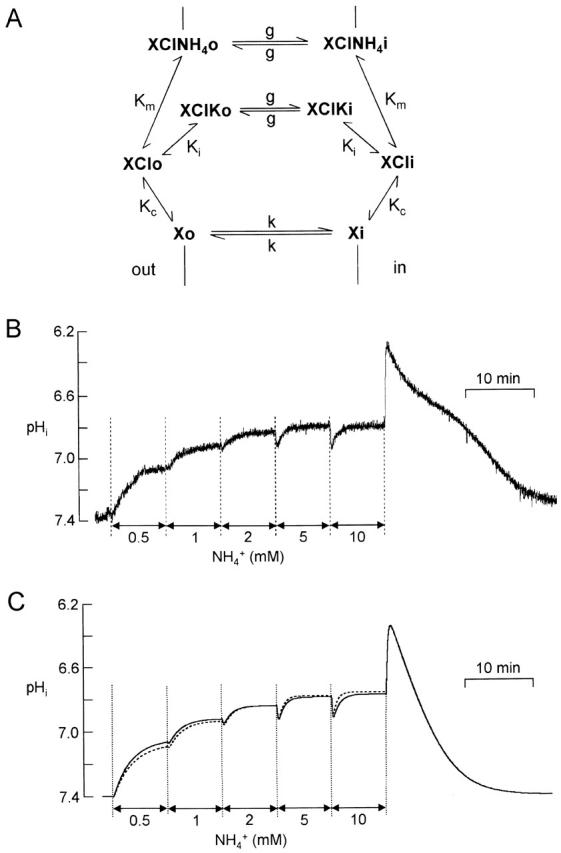
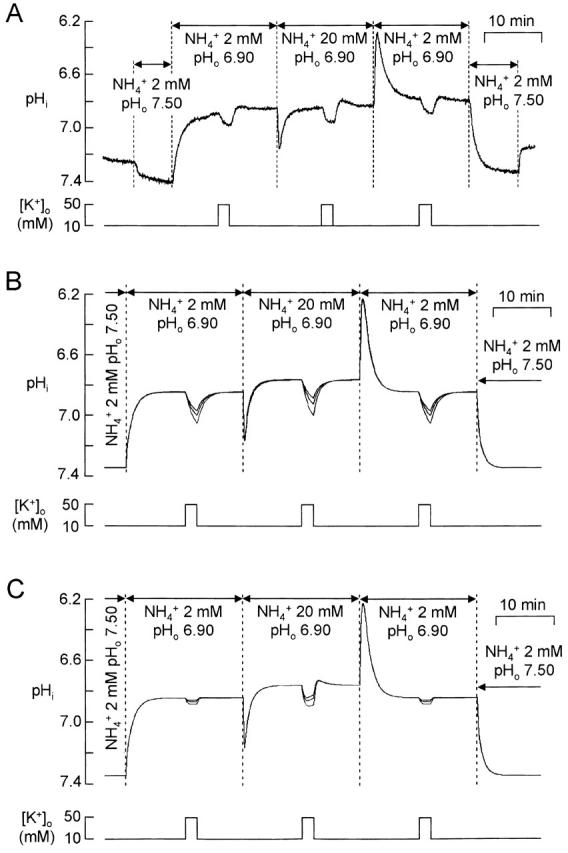
There appears to be a flux of ammonium (NH(4)(+)/NH(3)) from neurons to glial cells in most nervous tissues. In bee retinal glial cells, NH(4)(+)/NH(3) uptake is at least partly by chloride-dependant transport of the ionic form NH(4)(+). Transmembrane transport of NH(4)(+) has been described previously on transporters on which NH(4)(+) replaces K(+), or, more rarely, Na(+) or H(+), but no transport system in animal cells has been shown to be selective for NH(4)(+) over these other ions. To see if the NH(4)(+)-Cl(-) cotransporter on bee retinal glial cells is selective for NH(4)(+) over K(+) we measured ammonium-induced changes in intracellular pH (pH(i)) in isolated bundles of glial cells using a fluorescent indicator. These changes in pH(i) result from transmembrane fluxes not only of NH(4)(+), but also of NH(3). To estimate transmembrane fluxes of NH(4)(+), it was necessary to measure several parameters. Intracellular pH buffering power was found to be 12 mM. Regulatory mechanisms tended to restore intracellular [H(+)] after its displacement with a time constant of 3 min. Membrane permeability to NH(3) was 13 microm s(-1). A numerical model was used to deduce the NH(4)(+) flux through the transporter that would account for the pH(i) changes induced by a 30-s application of ammonium. This flux saturated with increasing [NH(4)(+)](o); the relation was fitted with a Michaelis-Menten equation with K(m) approximately 7 mM. The inhibition of NH(4)(+) flux by extracellular K(+) appeared to be competitive, with an apparent K(i) of approximately 15 mM. A simple standard model of the transport process satisfactorily described the pH(i) changes caused by various experimental manipulations when the transporter bound NH(4)(+) with greater affinity than K(+). We conclude that this transporter is functionally selective for NH(4)(+) over K(+) and that the transporter molecule probably has a greater affinity for NH(4)(+) than for K(+).
Figures
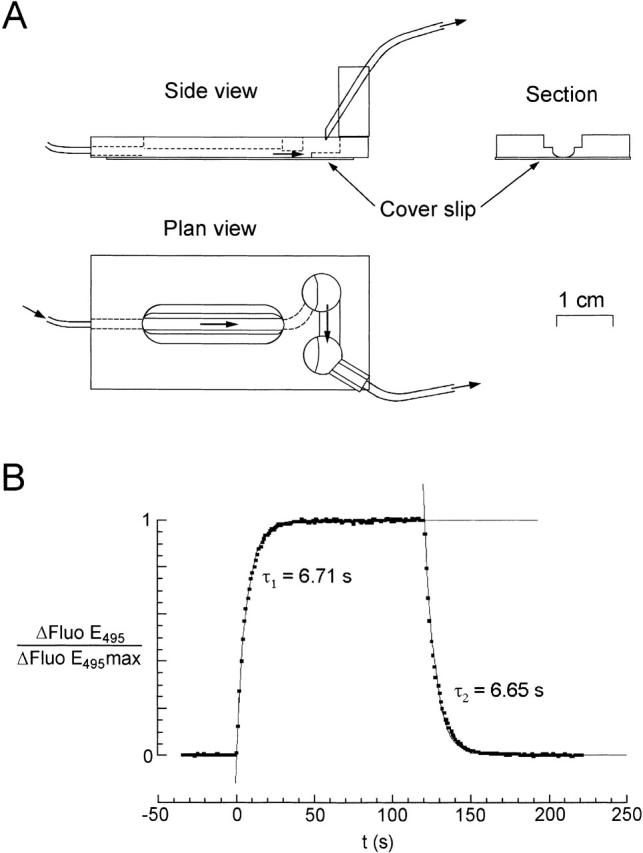
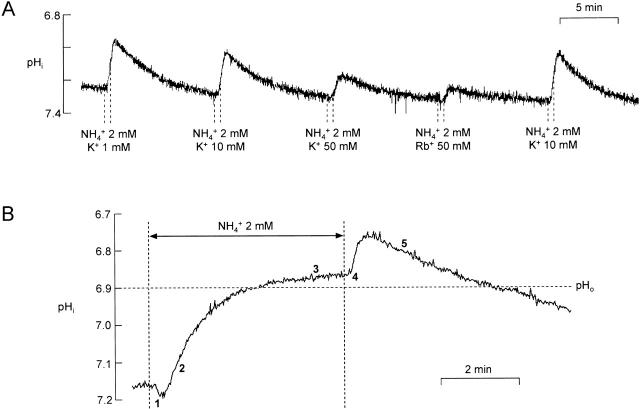
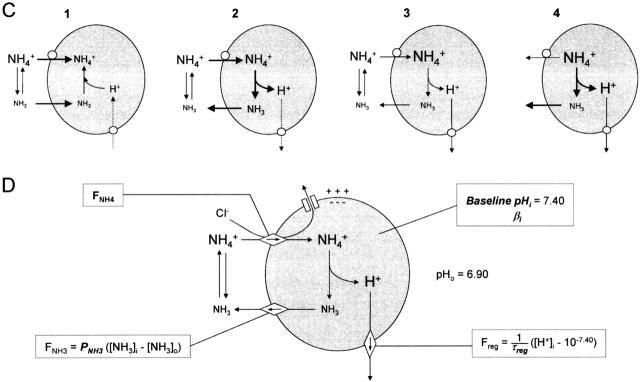
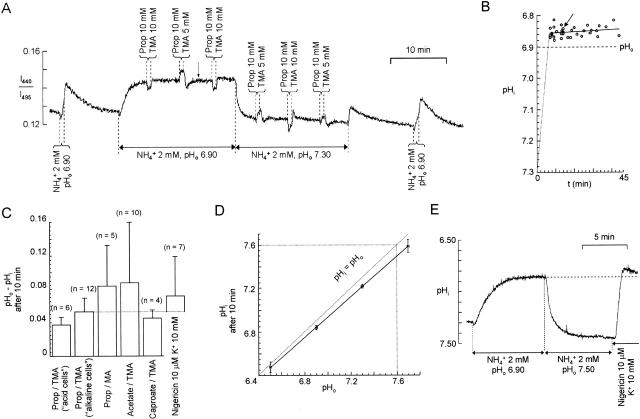
References
-
- Abramowitz M., Stegun I.A. Handbook of Mathematical Functions 1965. Dover Publications Inc; New York, NY: pp. 1046 pp
-
- Benjamin A.M., Quastel J.H. Metabolism of amino acids and ammonia in rat brain cortex slices in vitroa possible role of ammonia in brain function. J. Neurochem. 1975;25:197–206. - PubMed
-
- Bertrand, D. 1974. Etude des propriétés électrophysiologiques des cellules pigmentaires de la rétine du faux-bourdon (Apis mellifera). Thesis No. 1650. Université de Genéve.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
