

APC(ste9/srw1) promotes degradation of mitotic cyclins in G(1) and is inhibited by cdc2 phosphorylation
- PMID: 10921876
- PMCID: PMC306614
- DOI: 10.1093/emboj/19.15.3945
APC(ste9/srw1) promotes degradation of mitotic cyclins in G(1) and is inhibited by cdc2 phosphorylation
Abstract
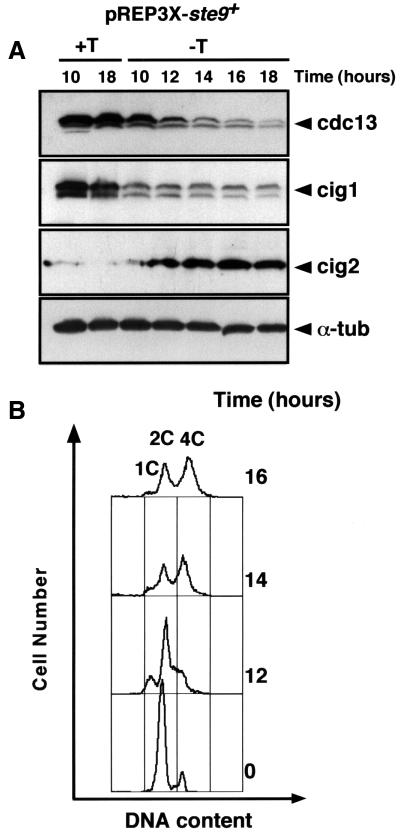
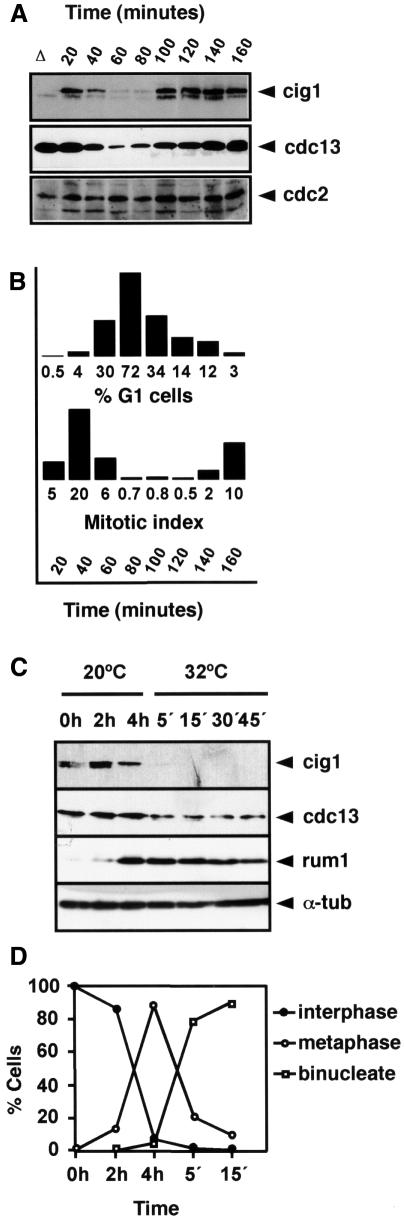
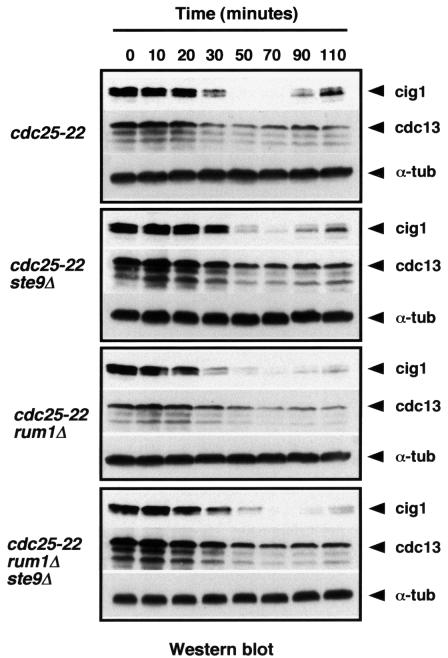
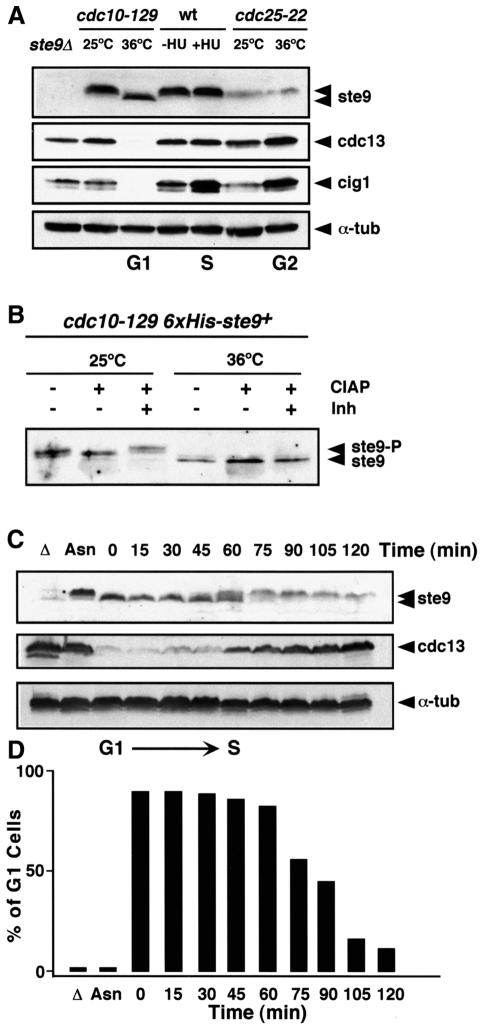
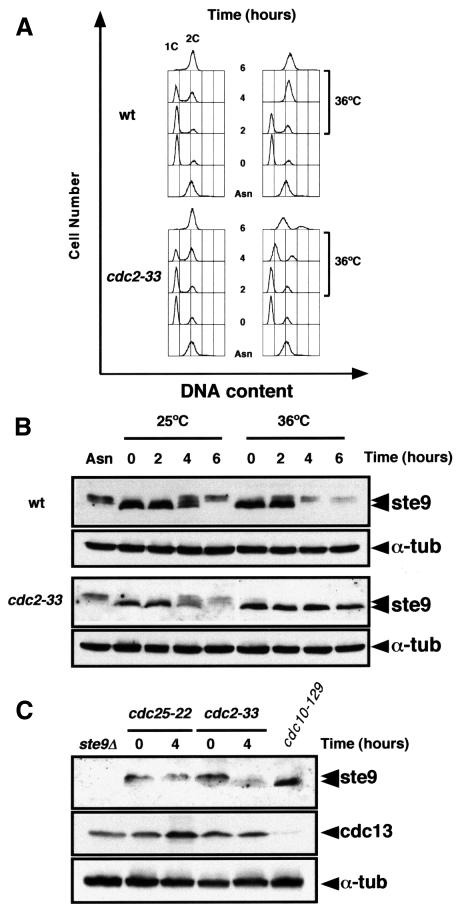
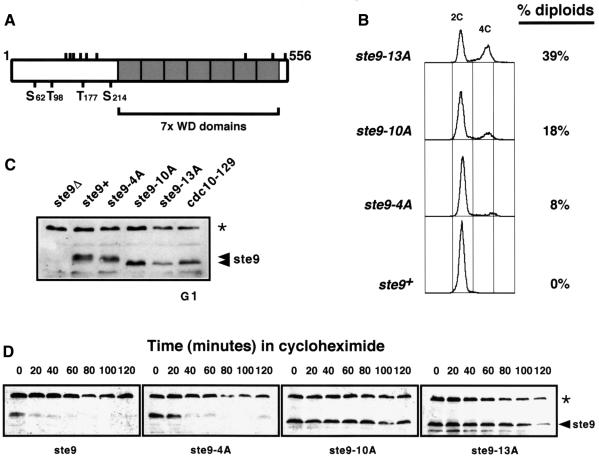
Fission yeast ste9/srw1 is a WD-repeat protein highly homologous to budding yeast Hct1/Cdh1 and Drosophila Fizzy-related that are involved in activating APC/C (anaphase-promoting complex/cyclosome). We show that APC(ste9/srw1) specifically promotes the degradation of mitotic cyclins cdc13 and cig1 but not the S-phase cyclin cig2. APC(ste9/srw1) is not necessary for the proteolysis of cdc13 and cig1 that occurs at the metaphase-anaphase transition but it is absolutely required for their degradation in G(1). Therefore, we propose that the main role of APC(ste9/srw1) is to promote degradation of mitotic cyclins when cells need to delay or arrest the cell cycle in G(1). We also show that ste9/srw1 is negatively regulated by cdc2-dependent protein phosphorylation. In G(1), when cdc2-cyclin kinase activity is low, unphosphorylated ste9/srw1 interacts with APC/C. In the rest of the cell cycle, phosphorylation of ste9/srw1 by cdc2-cyclin complexes both triggers proteolysis of ste9/srw1 and causes its dissociation from the APC/C. This mechanism provides a molecular switch to prevent inactivation of cdc2 in G(2) and early mitosis and to allow its inactivation in G(1).
Figures

References
-
- Amon A., Irniger,S. and Nasmyth,K. (1994) Closing the cell cycle circle in yeast: G2 cyclin proteolysis initiated at mitosis persits until the activation of G1 cyclins in the next cycle. Cell, 77, 1037–1050. - PubMed
-
- Baümer M., Braus,G.H. and Irniger,S. (2000) Two different modes of cyclin Clb2 proteolysis during mitosis in Saccharomyces cerevisae. FEBS Lett., 468, 142–148. - PubMed
-
- Boeke J.D., LaCroute,F. and Fink,G.R. (1984) A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol. Gen. Genet., 197, 345–346. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
