

The RET proto-oncogene induces apoptosis: a novel mechanism for Hirschsprung disease
- PMID: 10921886
- PMCID: PMC306592
- DOI: 10.1093/emboj/19.15.4056
The RET proto-oncogene induces apoptosis: a novel mechanism for Hirschsprung disease
Abstract
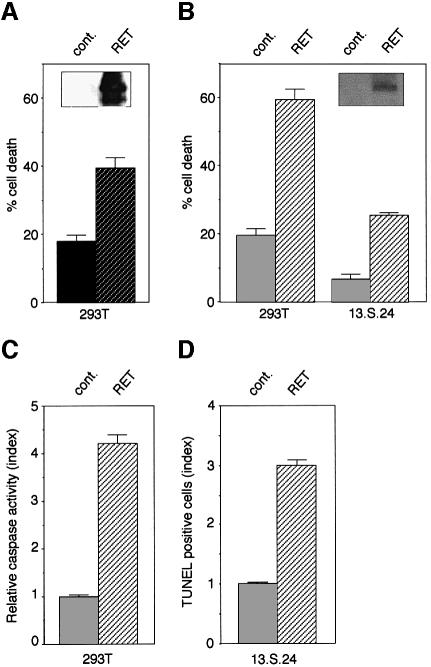
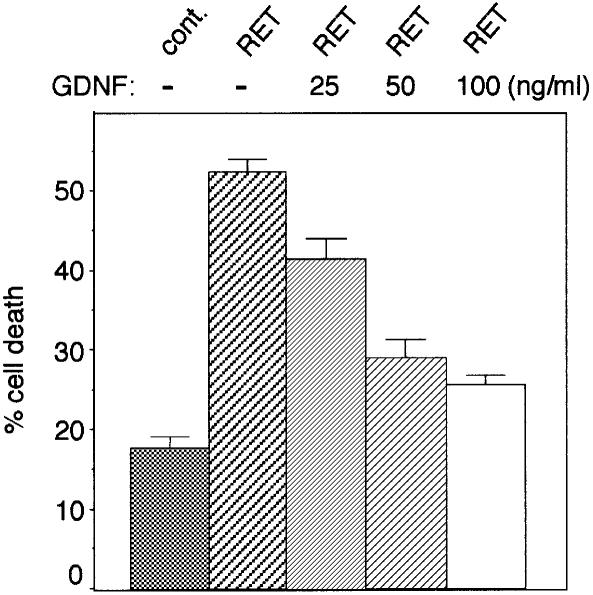
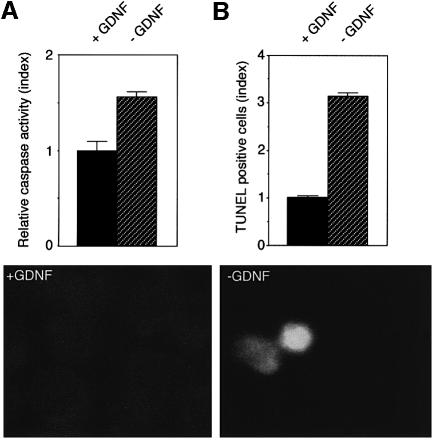
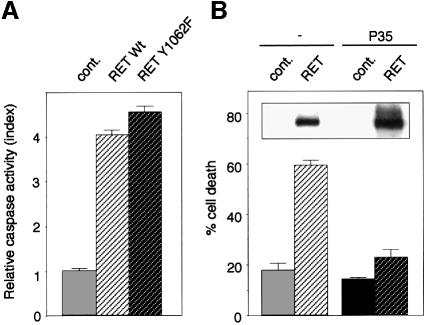
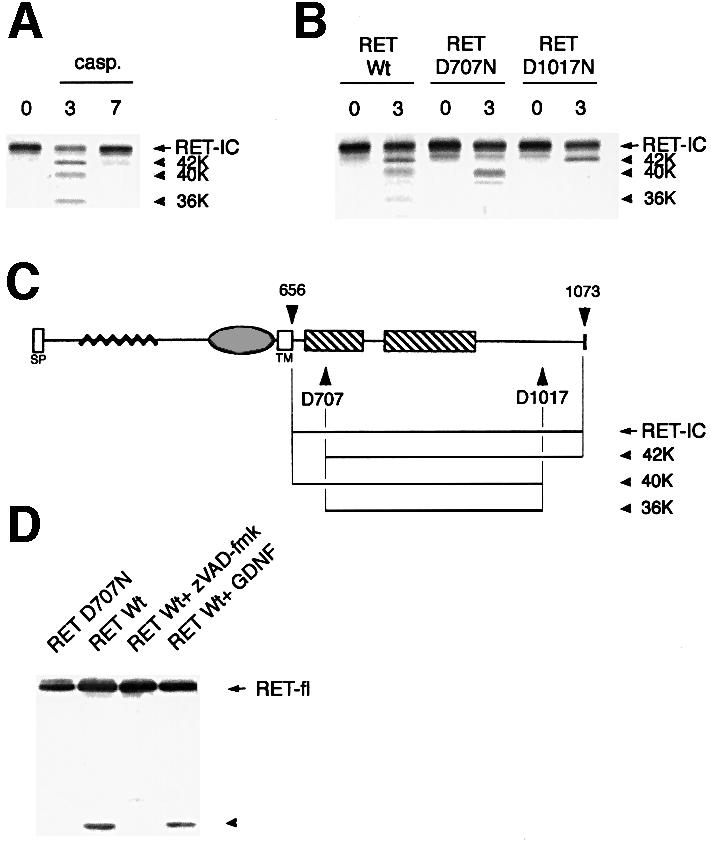
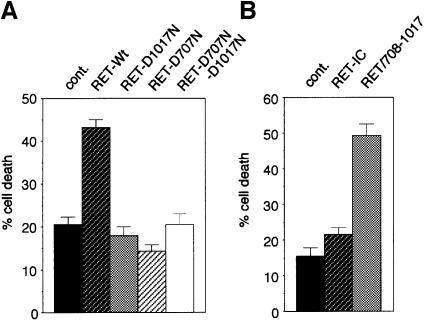
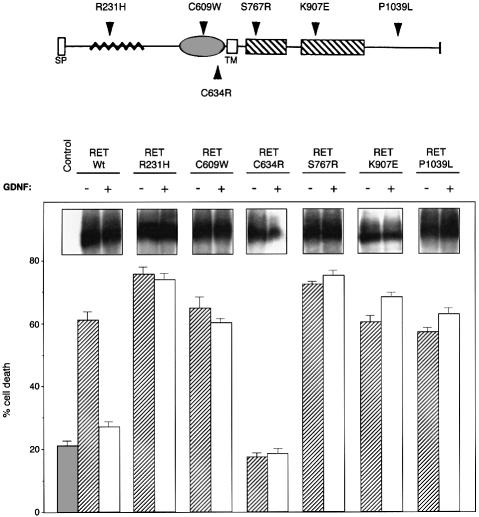
The RET (rearranged during transfection) proto-oncogene encodes a tyrosine kinase receptor involved in both multiple endocrine neoplasia type 2 (MEN 2), an inherited cancer syndrome, and Hirschsprung disease (HSCR), a developmental defect of enteric neurons. We report here that the expression of RET receptor induces apoptosis. This pro-apoptotic effect of RET is inhibited in the presence of its ligand glial cell line-derived neurotrophic factor (GDNF). Furthermore, we present evidence that RET induces apoptosis via its own cleavage by caspases, a phenomenon allowing the liberation/exposure of a pro-apoptotic domain of RET. In addition, we report that Hirschsprung-associated RET mutations impair GDNF control of RET pro-apoptotic activity. These results indicate that HSCR may result from apoptosis of RET-expressing enteric neuroblasts.
Figures
References
-
- Arighi E. et al. (1997) Identification of Shc docking site on Ret tyrosine kinase. Oncogene, 14, 773–782. - PubMed
-
- Baloh R.H. et al. (1997) TrnR2, a novel receptor that mediates neurturin and GDNF signaling through Ret. Neuron, 18, 793–802. - PubMed
-
- Bredesen D.E., Ye,X., Tasinato,A., Sperandio,S., Assa-Munt,N. and Rabizadeh,S. (1998) p75NTR and the concept of cellular dependence: seeing how the other half die. Cell Death Differ., 5, 365–371. - PubMed
-
- Cardone M.H., Roy,N., Stennicke,H.R., Salvesen,G.S., Franke,T.F., Stanbridge,E., Frisch,S. and Reed,J.C. (1998) Regulation of cell death protease caspase-9 by phosphorylation. Science, 282, 1318–1321. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
