

Role of the cystic fibrosis transmembrane conductance regulator in innate immunity to Pseudomonas aeruginosa infections
- PMID: 10922041
- PMCID: PMC34018
- DOI: 10.1073/pnas.97.16.8822
Role of the cystic fibrosis transmembrane conductance regulator in innate immunity to Pseudomonas aeruginosa infections
Abstract
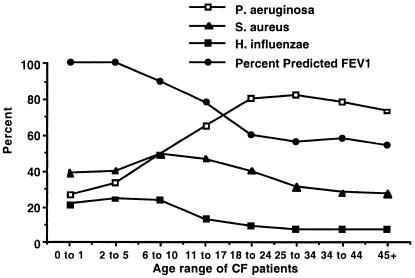
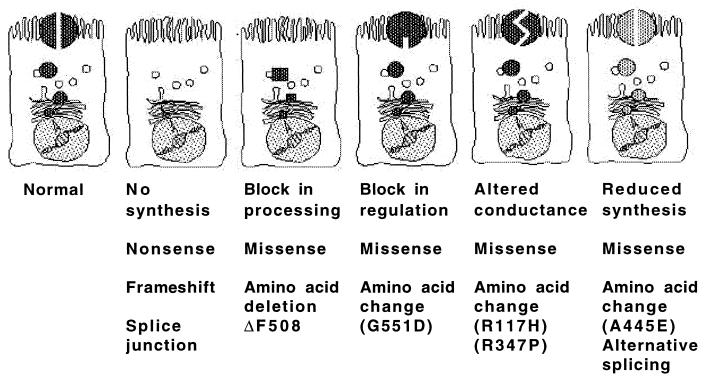
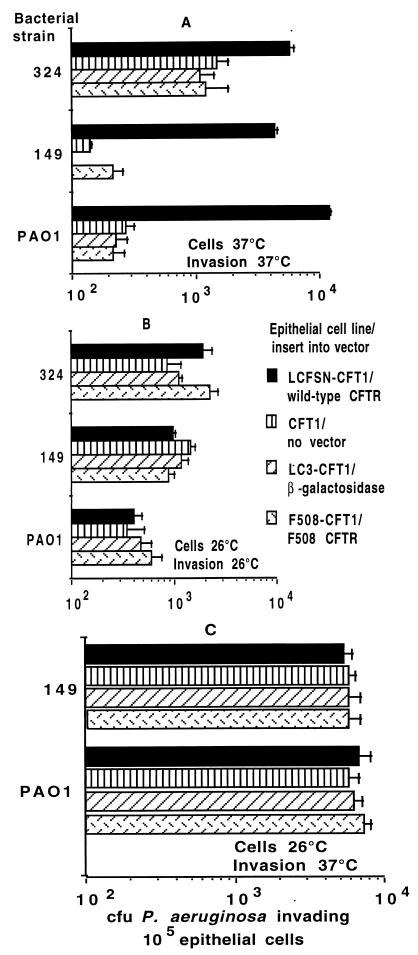
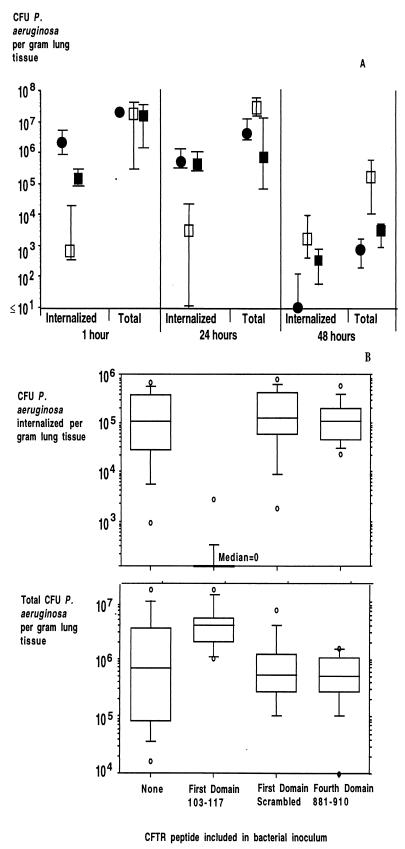
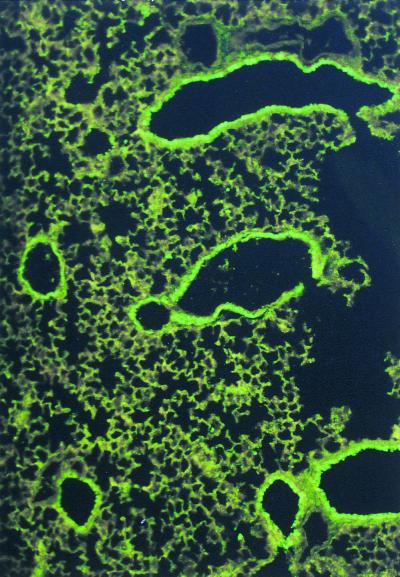
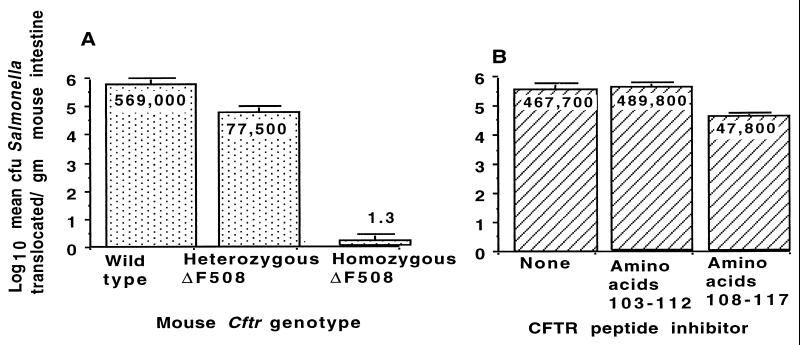
Chronic Pseudomonas aeruginosa infection occurs in 75-90% of patients with cystic fibrosis (CF). It is the foremost factor in pulmonary function decline and early mortality. A connection has been made between mutant or missing CF transmembrane conductance regulator (CFTR) in lung epithelial cell membranes and a failure in innate immunity leading to initiation of P. aeruginosa infection. Epithelial cells use CFTR as a receptor for internalization of P. aeruginosa via endocytosis and subsequent removal of bacteria from the airway. In the absence of functional CFTR, this interaction does not occur, allowing for increased bacterial loads in the lungs. Binding occurs between the outer core of the bacterial lipopolysaccharide and amino acids 108-117 in the first predicted extracellular domain of CFTR. In experimentally infected mice, inhibiting CFTR-mediated endocytosis of P. aeruginosa by inclusion in the bacterial inoculum of either free bacterial lipopolysaccharide or CFTR peptide 108-117 resulted in increased bacterial counts in the lungs. CFTR is also a receptor on gastrointestinal epithelial cells for Salmonella enterica serovar Typhi, the etiologic agent of typhoid fever. There was a significant decrease in translocation of this organism to the gastrointestinal submucosa in transgenic mice that are heterozygous carriers of a mutant DeltaF508 CFTR allele, suggesting heterozygous CFTR carriers may have increased resistance to typhoid fever. The identification of CFTR as a receptor for bacterial pathogens could underlie the biology of CF lung disease and be the basis for the heterozygote advantage for carriers of mutant alleles of CFTR.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
