

Nitric oxide and salicylic acid signaling in plant defense
- PMID: 10922045
- PMCID: PMC34022
- DOI: 10.1073/pnas.97.16.8849
Nitric oxide and salicylic acid signaling in plant defense
Abstract
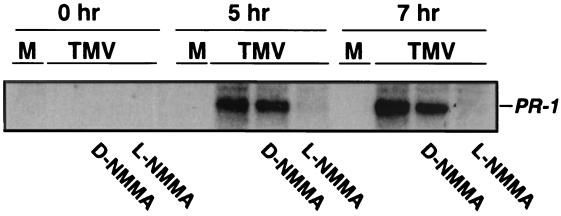
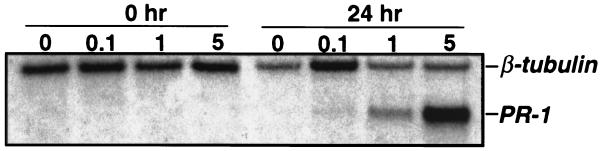
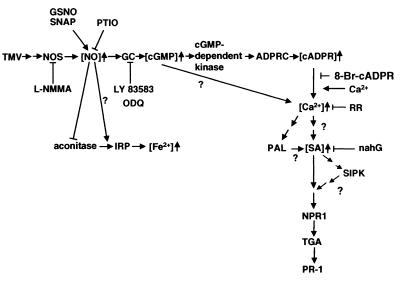
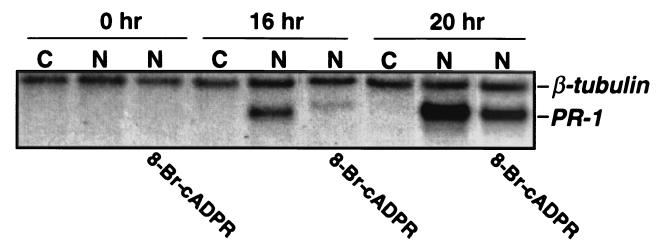
Salicylic acid (SA) plays a critical signaling role in the activation of plant defense responses after pathogen attack. We have identified several potential components of the SA signaling pathway, including (i) the H(2)O(2)-scavenging enzymes catalase and ascorbate peroxidase, (ii) a high affinity SA-binding protein (SABP2), (iii) a SA-inducible protein kinase (SIPK), (iv) NPR1, an ankyrin repeat-containing protein that exhibits limited homology to IkappaBalpha and is required for SA signaling, and (v) members of the TGA/OBF family of bZIP transcription factors. These bZIP factors physically interact with NPR1 and bind the SA-responsive element in promoters of several defense genes, such as the pathogenesis-related 1 gene (PR-1). Recent studies have demonstrated that nitric oxide (NO) is another signal that activates defense responses after pathogen attack. NO has been shown to play a critical role in the activation of innate immune and inflammatory responses in animals. Increases in NO synthase (NOS)-like activity occurred in resistant but not susceptible tobacco after infection with tobacco mosaic virus. Here we demonstrate that this increase in activity participates in PR-1 gene induction. Two signaling molecules, cGMP and cyclic ADP ribose (cADPR), which function downstream of NO in animals, also appear to mediate plant defense gene activation (e.g., PR-1). Additionally, NO may activate PR-1 expression via an NO-dependent, cADPR-independent pathway. Several targets of NO in animals, including guanylate cyclase, aconitase, and mitogen-activated protein kinases (e.g., SIPK), are also modulated by NO in plants. Thus, at least portions of NO signaling pathways appear to be shared between plants and animals.
Figures
Similar articles
-
Differential induction of tobacco MAP kinases by the defense signals nitric oxide, salicylic acid, ethylene, and jasmonic acid.Mol Plant Microbe Interact. 2000 Mar;13(3):347-51. doi: 10.1094/MPMI.2000.13.3.347. Mol Plant Microbe Interact. 2000. PMID: 10707361
-
Elucidation of signaling molecules involved in ergosterol perception in tobacco.Plant Physiol Biochem. 2013 Dec;73:121-7. doi: 10.1016/j.plaphy.2013.09.009. Epub 2013 Sep 20. Plant Physiol Biochem. 2013. PMID: 24095918
-
LW-1 induced resistance to TMV in tobacco was mediated by nitric oxide and salicylic acid pathway.Pestic Biochem Physiol. 2024 Jun;202:105896. doi: 10.1016/j.pestbp.2024.105896. Epub 2024 Apr 4. Pestic Biochem Physiol. 2024. PMID: 38879345
-
NPR1: the spider in the web of induced resistance signaling pathways.Curr Opin Plant Biol. 2004 Aug;7(4):456-64. doi: 10.1016/j.pbi.2004.05.006. Curr Opin Plant Biol. 2004. PMID: 15231270 Review.
-
Pandemonium Breaks Out: Disruption of Salicylic Acid-Mediated Defense by Plant Pathogens.Mol Plant. 2018 Dec 3;11(12):1427-1439. doi: 10.1016/j.molp.2018.10.002. Epub 2018 Oct 15. Mol Plant. 2018. PMID: 30336330 Review.
Cited by
-
Characterization of secretory phospholipase A₂ with phospholipase A₁ activity in tobacco, Nicotiana tabacum (L.).Lipids. 2012 Mar;47(3):303-12. doi: 10.1007/s11745-011-3632-3. Epub 2011 Nov 29. Lipids. 2012. PMID: 22124805
-
The structure of a major surface antigen SAG19 from Eimeria tenella unifies the Eimeria SAG family.Commun Biol. 2021 Mar 19;4(1):376. doi: 10.1038/s42003-021-01904-w. Commun Biol. 2021. PMID: 33742128 Free PMC article.
-
Constitutive expression of mammalian nitric oxide synthase in tobacco plants triggers disease resistance to pathogens.Mol Cells. 2012 Nov;34(5):463-71. doi: 10.1007/s10059-012-0213-0. Epub 2012 Oct 31. Mol Cells. 2012. PMID: 23124383 Free PMC article.
-
Structural studies of soybean calmodulin isoform 4 bound to the calmodulin-binding domain of tobacco mitogen-activated protein kinase phosphatase-1 provide insights into a sequential target binding mode.J Biol Chem. 2009 Oct 9;284(41):28292-28305. doi: 10.1074/jbc.M109.025080. Epub 2009 Aug 10. J Biol Chem. 2009. PMID: 19667066 Free PMC article.
-
Reactive oxygen species and reactive nitrogen species in peroxisomes. Production, scavenging, and role in cell signaling.Plant Physiol. 2006 Jun;141(2):330-5. doi: 10.1104/pp.106.078204. Plant Physiol. 2006. PMID: 16760483 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
