

The junctional multidomain protein AF-6 is a binding partner of the Rap1A GTPase and associates with the actin cytoskeletal regulator profilin
- PMID: 10922060
- PMCID: PMC16822
- DOI: 10.1073/pnas.97.16.9064
The junctional multidomain protein AF-6 is a binding partner of the Rap1A GTPase and associates with the actin cytoskeletal regulator profilin
Abstract
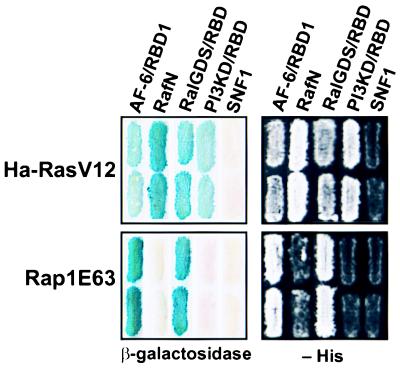
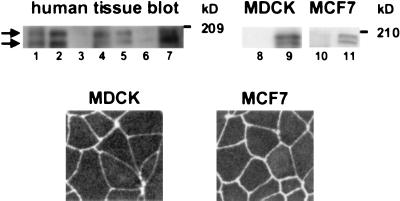
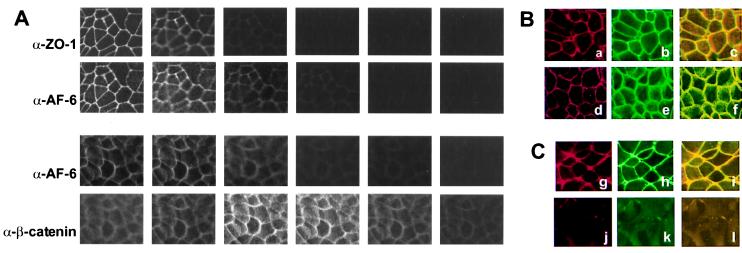
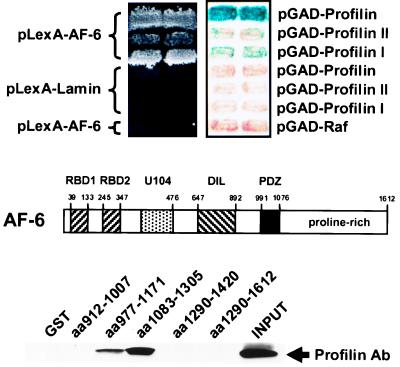
The AF-6 protein is a multidomain protein that contains two potential Ras-binding domains within its N terminus. Because of this feature, AF-6 has been isolated in both two-hybrid and biochemical approaches and is postulated to be a potential Ras-effector protein. Herein, we show that it is specifically the first Ras-binding domain of AF-6 that mediates this interaction and that the Ras-related Rap1A protein can associate with this motif even more efficiently than the oncogenic Ha-, K-, and N-Ras GTPases. We further demonstrate that both Ras and Rap1 interact with full-length AF-6 in vivo in mammalian cells and that a fraction of Rap1 colocalizes with AF-6 at the membrane. Dominant active Rap1A, in contrast to Ras, when introduced into epithelial MDCK and MCF-7 cells, does not perturb AF-6-specific residency in cell-cell adhesion complexes. In a pursuit to gain further understanding of the role of AF-6 in junctions, we identified profilin as an AF-6-binding protein. Profilin activates monomeric actin units for subsequent polymerization steps at barbed ends of actin filaments and has been shown to participate in cortical actin assembly. To our knowledge, AF-6 is the only integral component in cell-cell junctions discovered thus far that interacts with profilin and thus could modulate actin modeling proximal to adhesion complexes.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
