

The yeast TEL1 gene partially substitutes for human ATM in suppressing hyperrecombination, radiation-induced apoptosis and telomere shortening in A-T cells
- PMID: 10930457
- PMCID: PMC14943
- DOI: 10.1091/mbc.11.8.2605
The yeast TEL1 gene partially substitutes for human ATM in suppressing hyperrecombination, radiation-induced apoptosis and telomere shortening in A-T cells
Abstract
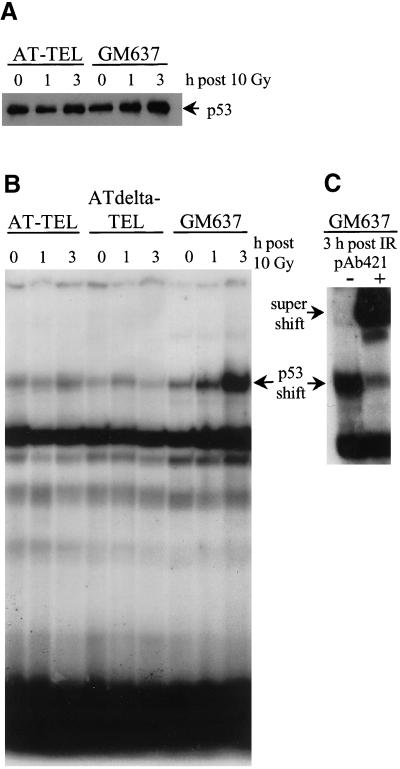
Homozygous mutations in the human ATM gene lead to a pleiotropic clinical phenotype of ataxia-telangiectasia (A-T) patients and correlating cellular deficiencies in cells derived from A-T donors. Saccharomyces cerevisiae tel1 mutants lacking Tel1p, which is the closest sequence homologue to the ATM protein, share some of the cellular defects with A-T. Through genetic complementation of A-T cells with the yeast TEL1 gene, we provide evidence that Tel1p can partially compensate for ATM in suppressing hyperrecombination, radiation-induced apoptosis, and telomere shortening. Complementation appears to be independent of p53 activation. The data provided suggest that TEL1 is a functional homologue of human ATM in yeast, and they help to elucidate different cellular and biochemical pathways in human cells regulated by the ATM protein.
Figures
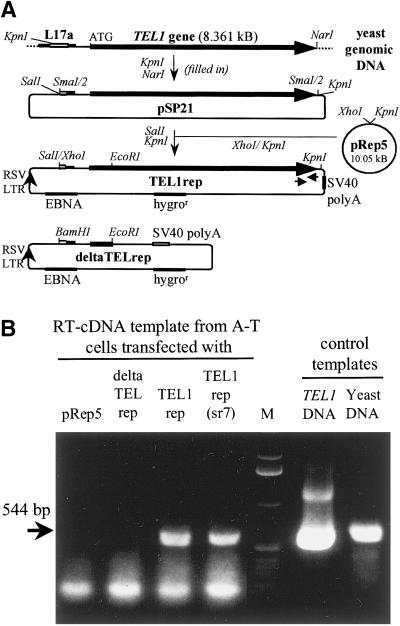
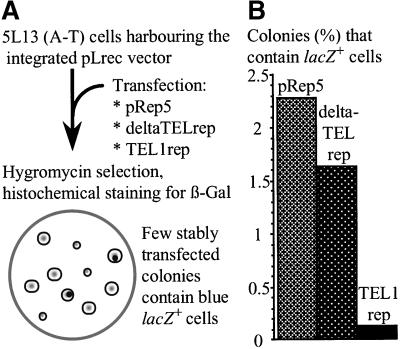
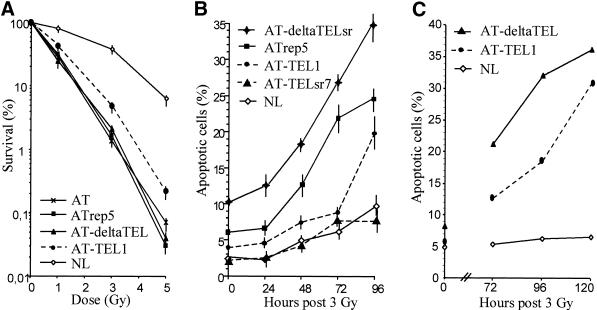
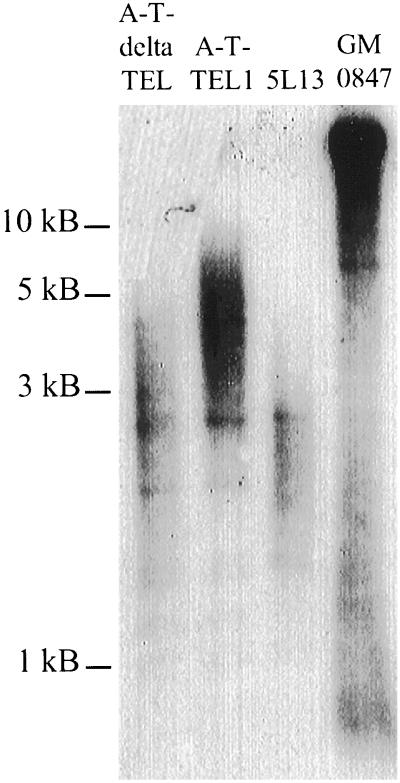
References
-
- Aurias A, Dutrillaux B, Buriot D, Lejeune J. High frequencies of inversions and translocations of chromosomes 7 and 14 in ataxia telangiectasia. Mutat Res. 1980;69:369–74. - PubMed
-
- Banin S, Moyal L, Shieh S, Taya Y, Anderson CW, Chessa L, Smorodinsky NI, Prives C, Reiss Y, Shiloh Y, Ziv Y. Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science. 1998;281:1674–1677. - PubMed
-
- Baskaran R, Wood LD, Whitaker LL, Canman CE, Morgan SE, Xu Y, Barlow C, Baltimore D, Wynshaw-Boris A, Kastan MB, Wang JY. Ataxia telangiectasia mutant protein activates c-Abl tyrosine kinase in response to ionizing radiation. Nature. 1997;387:516–519. - PubMed
-
- Beamish H, Lavin MF. Radiosensitivity in ataxia-telangiectasia: anomalies in radiation-induced cell cycle delay. Int J Radiat Biol. 1994;65:175–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
