

The interaction between Ran and NTF2 is required for cell cycle progression
- PMID: 10930458
- PMCID: PMC14944
- DOI: 10.1091/mbc.11.8.2617
The interaction between Ran and NTF2 is required for cell cycle progression
Abstract
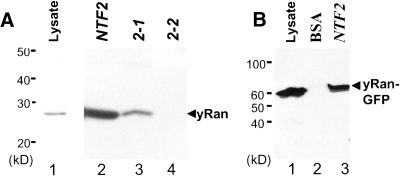
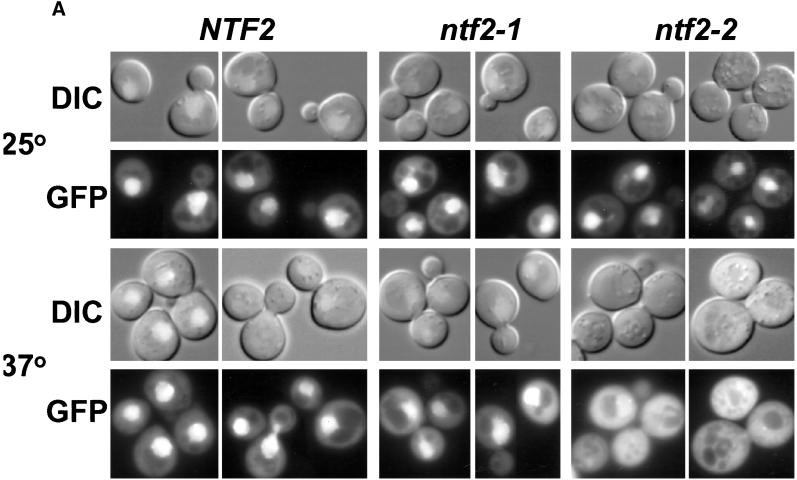
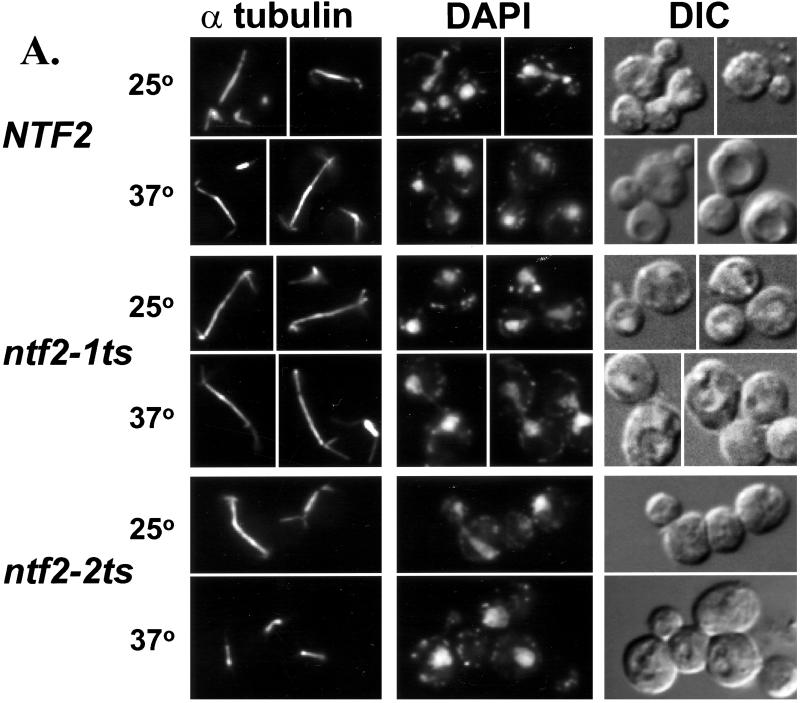
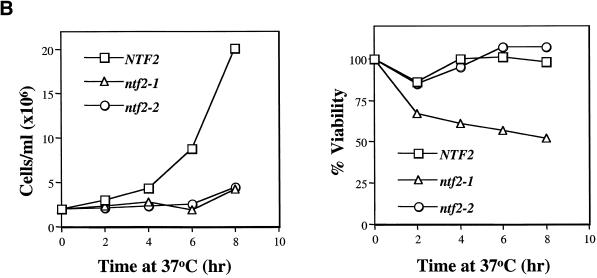
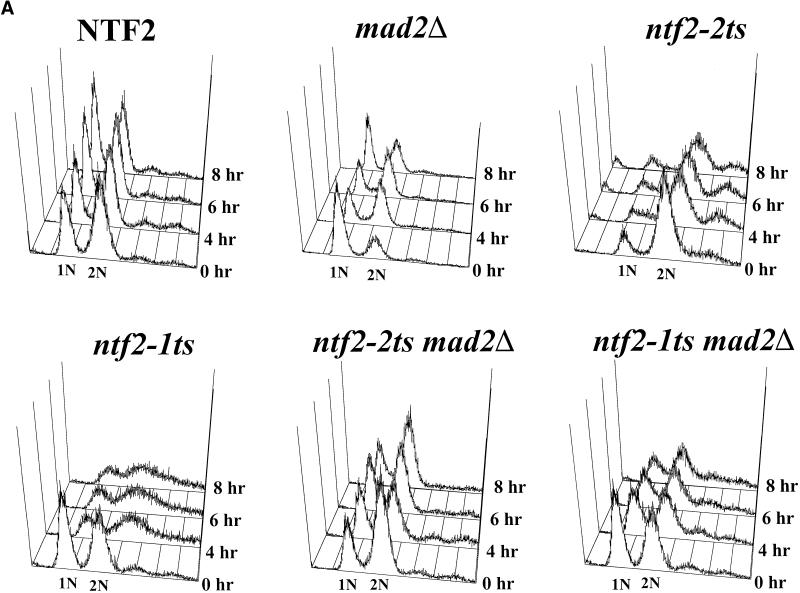
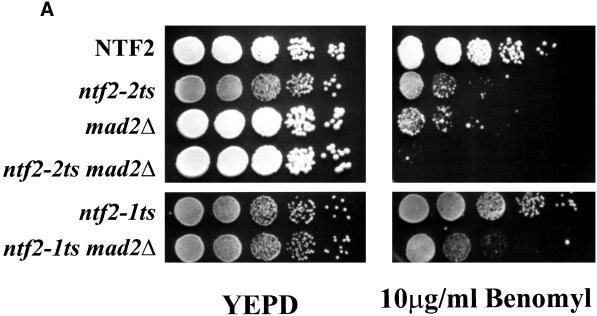
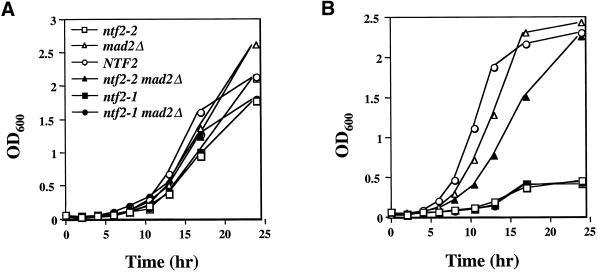
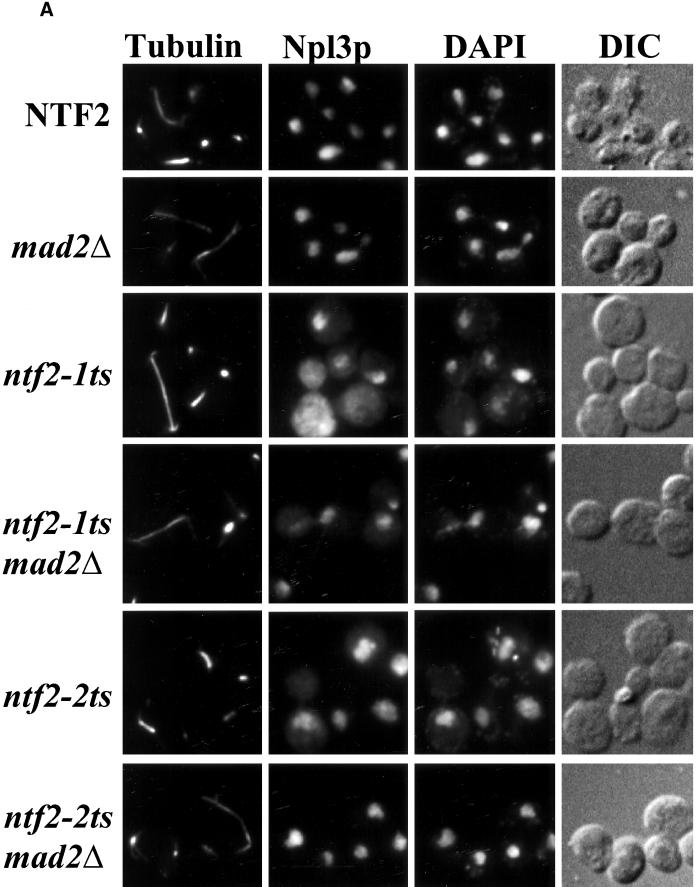
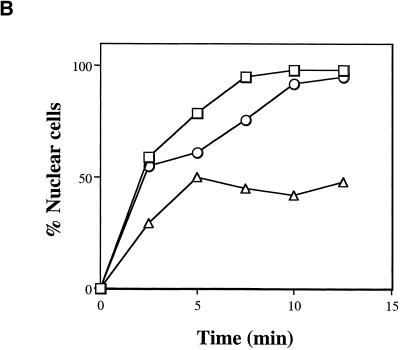
The small GTPase Ran is required for the trafficking of macromolecules into and out of the nucleus. Ran also has been implicated in cell cycle control, specifically in mitotic spindle assembly. In interphase cells, Ran is predominately nuclear and thought to be GTP bound, but it is also present in the cytoplasm, probably in the GDP-bound state. Nuclear transport factor 2 (NTF2) has been shown to import RanGDP into the nucleus. Here, we examine the in vivo role of NTF2 in Ran import and the effect that disruption of Ran imported into the nucleus has on the cell cycle. A temperature-sensitive (ts) mutant of Saccharomyces cerevisiae NTF2 that does not bind to Ran is unable to import Ran into the nucleus at the nonpermissive temperature. Moreover, when Ran is inefficiently imported into the nucleus, cells arrest in G(2) in a MAD2 checkpoint-dependent manner. These findings demonstrate that NTF2 is required to transport Ran into the nucleus in vivo. Furthermore, we present data that suggest that depletion of nuclear Ran triggers a spindle-assembly checkpoint-dependent cell cycle arrest.
Figures


References
-
- Adams A, Gottschling DE, Kaiser CA, Stearns T. Methods in Yeast Genetics. Cold Spring Harbor NY: Cold Spring Harbor Laboratory Press; 1997.
-
- Bischoff FR, Ponstingl H. Catalysis of guanine nucleotide exchange on Ran by the mitotic regulator RCC1. Nature. 1991;354:80–82. - PubMed
-
- Bullock TL, Clarkson WD, Kent HM, Stewart M. The 1.6Å resolution crystal structure of nuclear transport factor 2 (NTF2) J Mol Biol. 1996;260:422–431. - PubMed
-
- Carazo-Salas RE, Guarguaglini G, Gruss OJ, Segref A, Karsenti E, Mattaj IW. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature. 1999;400:178–181. - PubMed
-
- Chalfie M, Tu Y, Euskirchen W, Ward W, Prasher DC. Green fluorescent protein as a marker for gene expression. Science. 1994;263:802–805. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
