

Three regions within ActA promote Arp2/3 complex-mediated actin nucleation and Listeria monocytogenes motility
- PMID: 10931865
- PMCID: PMC2175181
- DOI: 10.1083/jcb.150.3.527
Three regions within ActA promote Arp2/3 complex-mediated actin nucleation and Listeria monocytogenes motility
Abstract
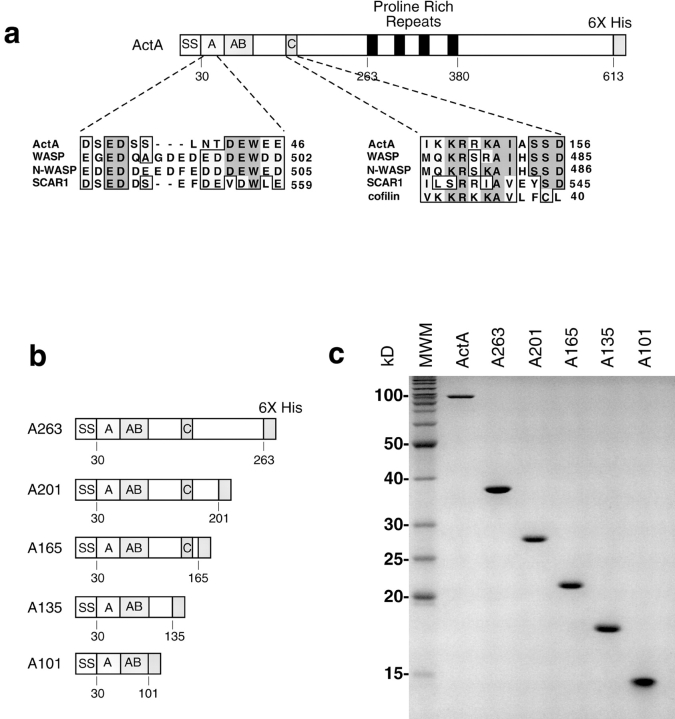
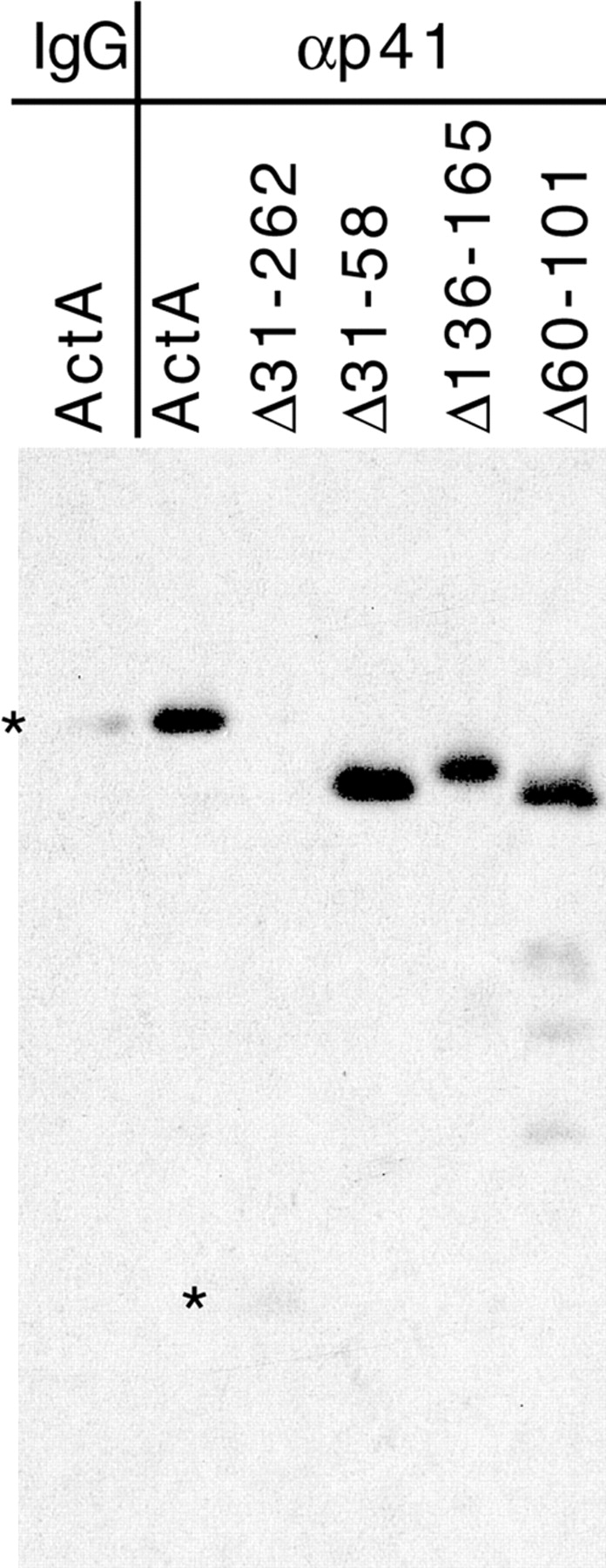
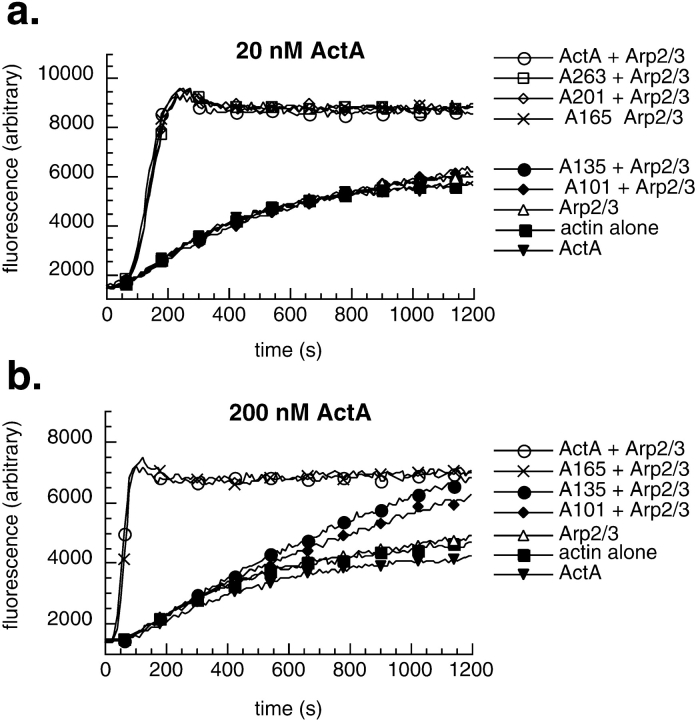
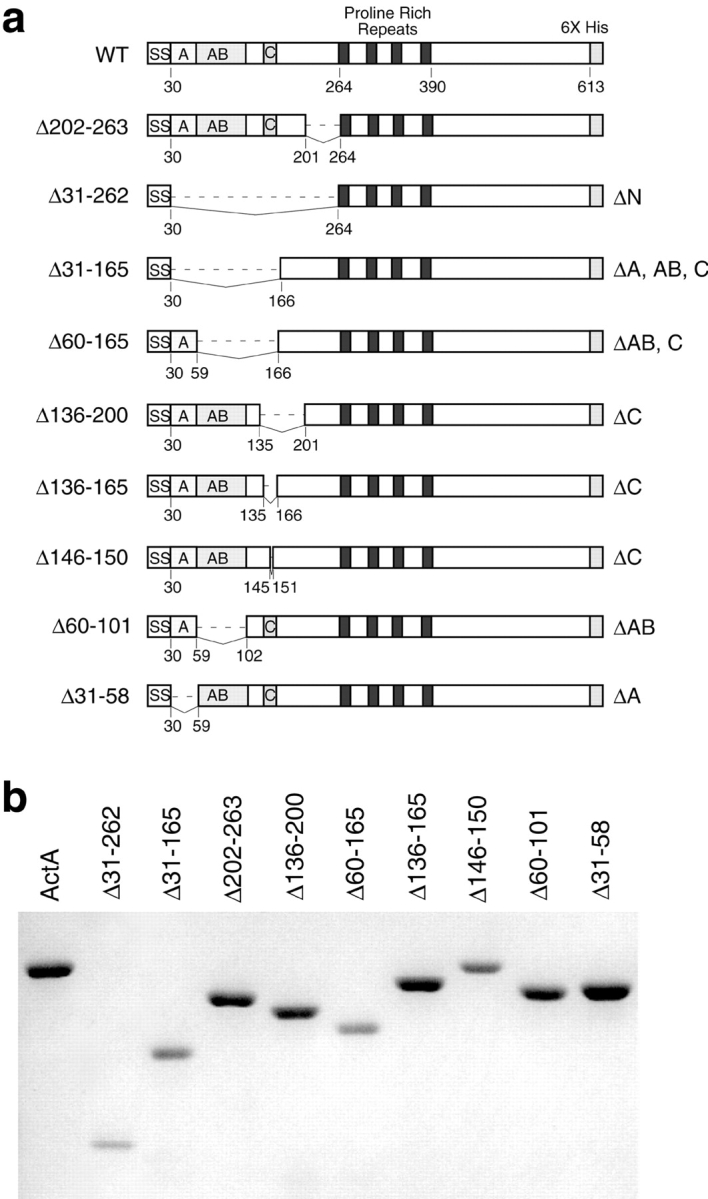
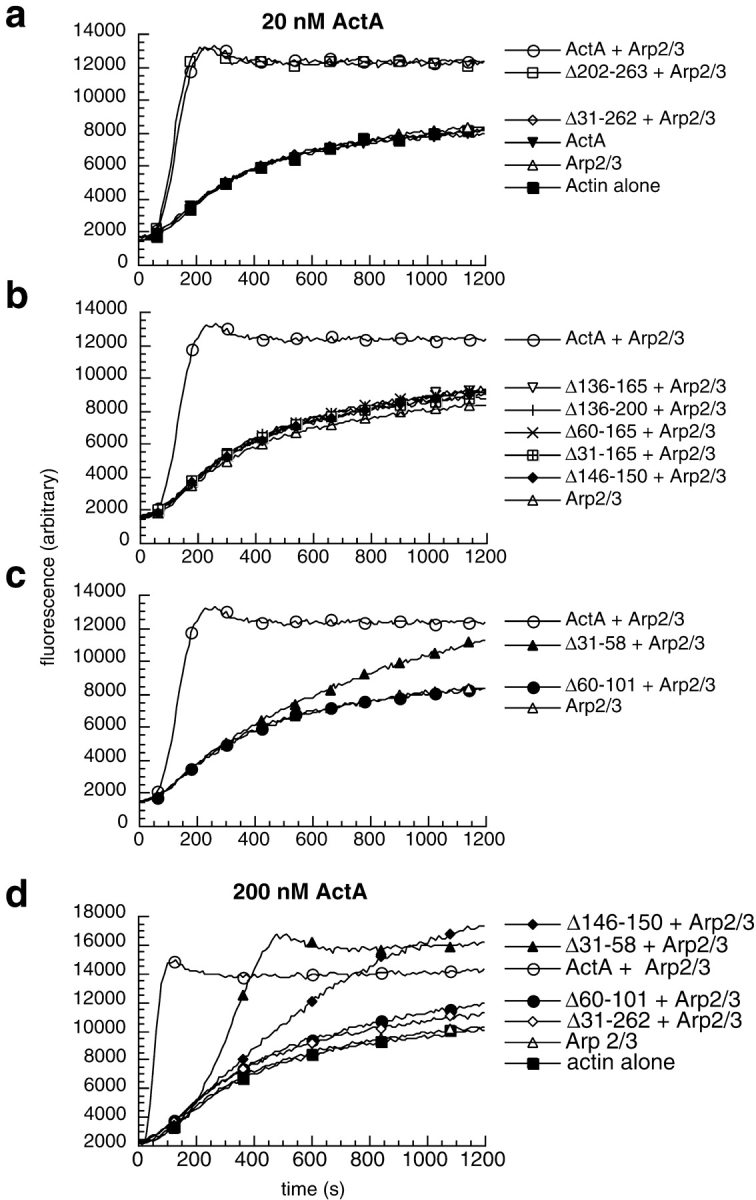
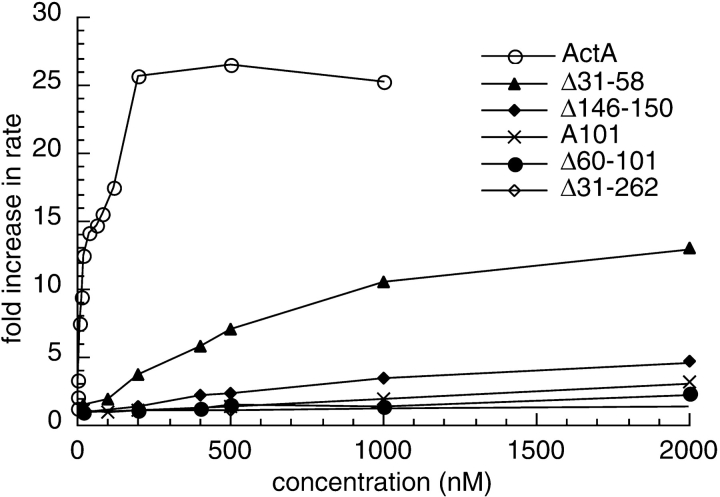
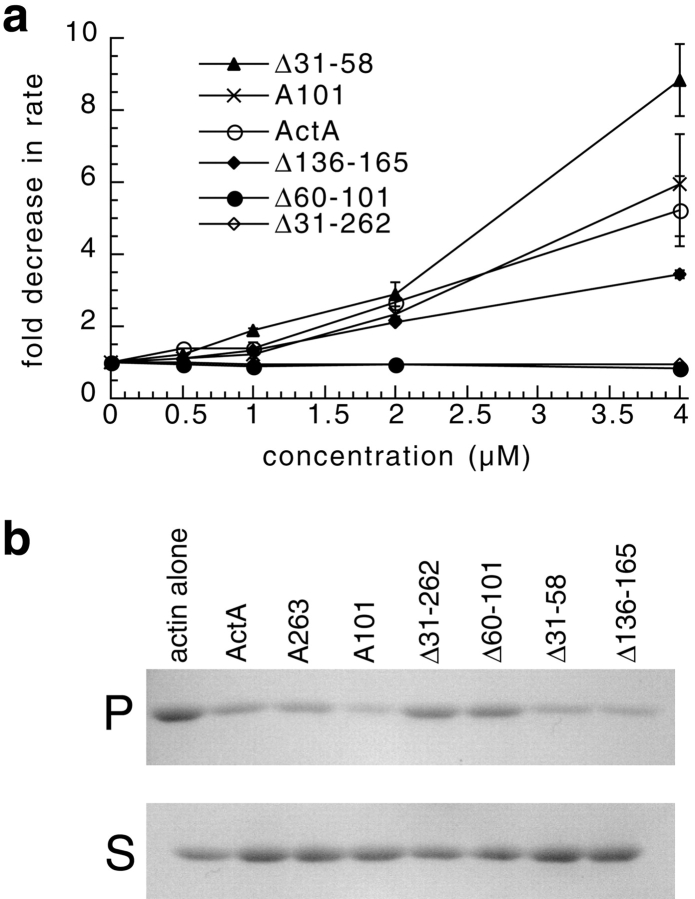
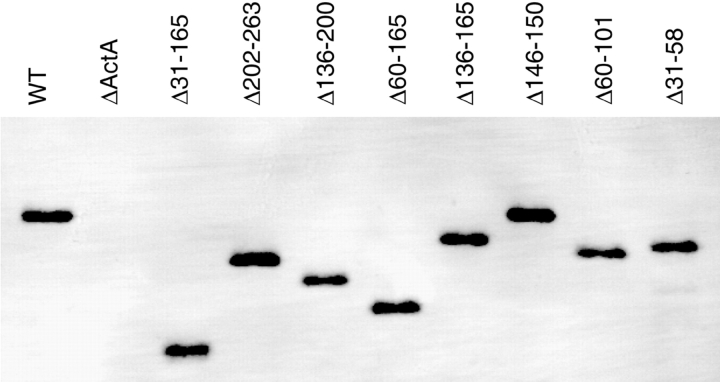
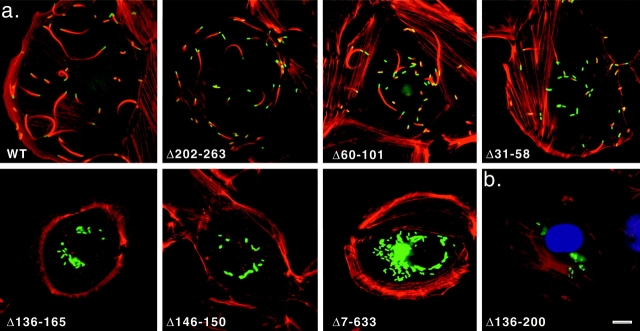
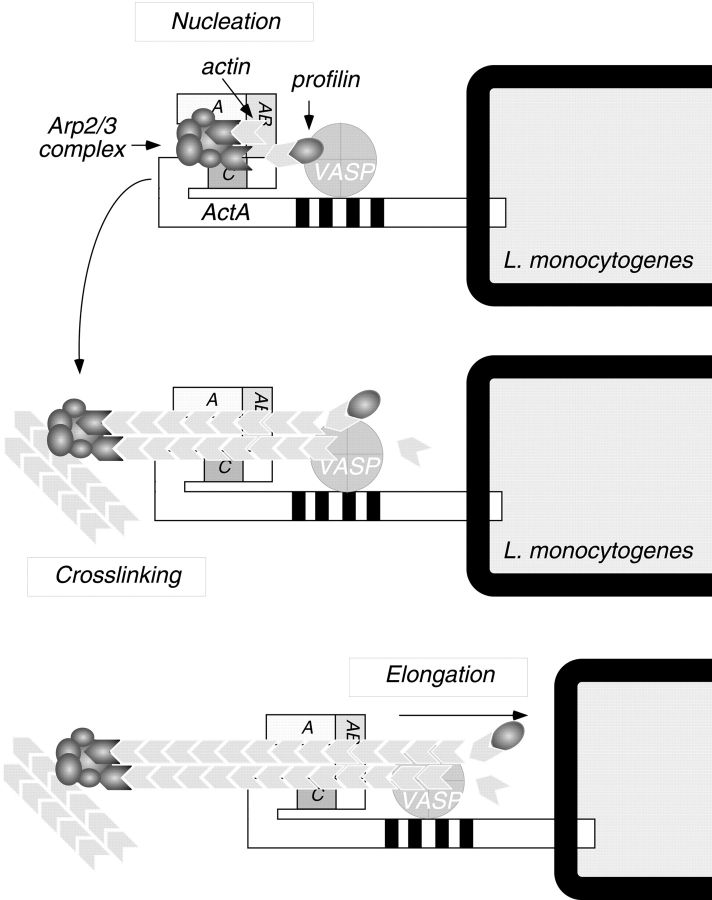
The Listeria monocytogenes ActA protein induces actin-based motility by enhancing the actin nucleating activity of the host Arp2/3 complex. Using systematic truncation analysis, we identified a 136-residue NH(2)-terminal fragment that was fully active in stimulating nucleation in vitro. Further deletion analysis demonstrated that this fragment contains three regions, which are important for nucleation and share functional and/or limited sequence similarity with host WASP family proteins: an acidic stretch, an actin monomer-binding region, and a cofilin homology sequence. To determine the contribution of each region to actin-based motility, we compared the biochemical activities of ActA derivatives with the phenotypes of corresponding mutant bacteria in cells. The acidic stretch functions to increase the efficiency of actin nucleation, the rate and frequency of motility, and the effectiveness of cell-cell spread. The monomer-binding region is required for actin nucleation in vitro, but not for actin polymerization or motility in infected cells, suggesting that redundant mechanisms may exist to recruit monomer in host cytosol. The cofilin homology sequence is critical for stimulating actin nucleation with the Arp2/3 complex in vitro, and is essential for actin polymerization and motility in cells. These data demonstrate that each region contributes to actin-based motility, and that the cofilin homology sequence plays a principal role in activation of the Arp2/3 complex, and is an essential determinant of L. monocytogenes pathogenesis.
Figures
References
-
- Banzai Y., Miki H., Yamaguchi H., Takenawa T. Essential role of neural Wiskott-Aldrich syndrome protein in neurite extension in PC12 cells and rat hippocampal primary culture cells. J. Biol. Chem. 2000;275:11987–11992. - PubMed
-
- Bachmann C., Fischer L., Walter U., Reinhard M. The EVH2 domain of the vasodilator-stimulated phosphoprotein mediates tetramerization, F-actin binding, and actin bundle formation. J. Biol. Chem. 1999;274:23549–23557. - PubMed
-
- Bi E., Zigmond S.H. Actin polymerizationwhere the WASP stings. Curr. Biol. 1999;9:R160–R163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
