

Regulation of the Epstein-Barr virus C promoter by AUF1 and the cyclic AMP/protein kinase A signaling pathway
- PMID: 10933728
- PMCID: PMC112351
- DOI: 10.1128/jvi.74.17.8166-8175.2000
Regulation of the Epstein-Barr virus C promoter by AUF1 and the cyclic AMP/protein kinase A signaling pathway
Abstract
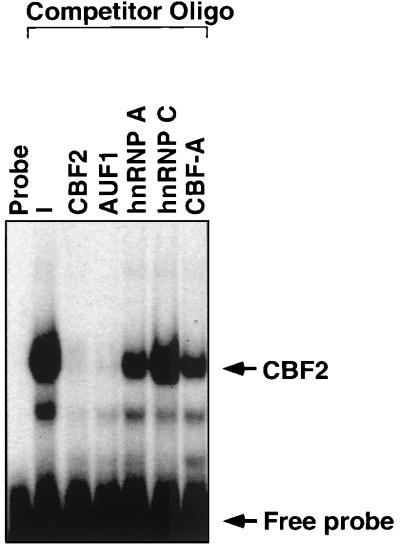
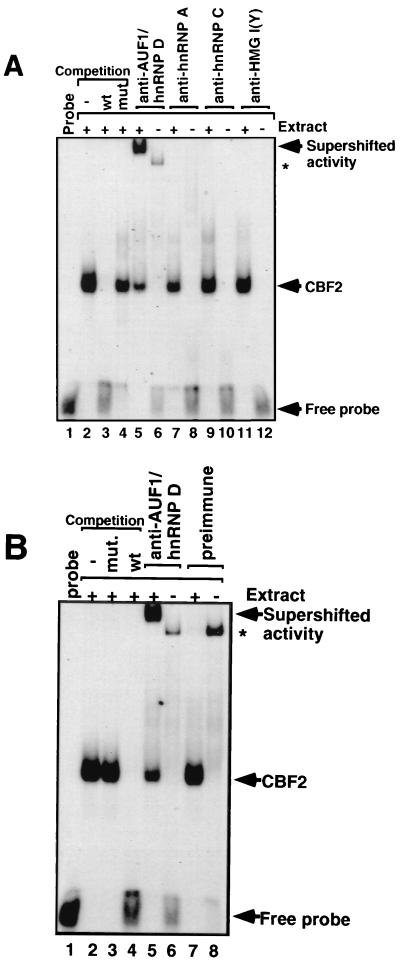
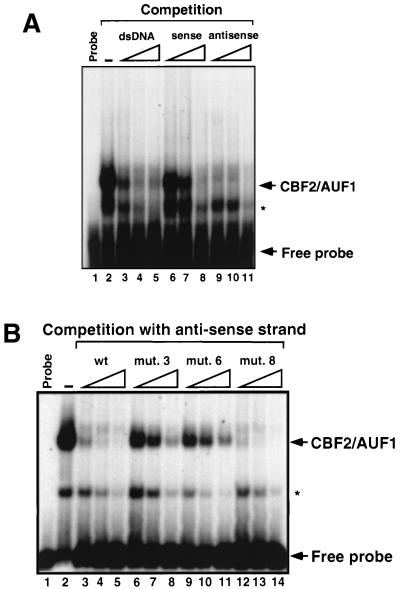
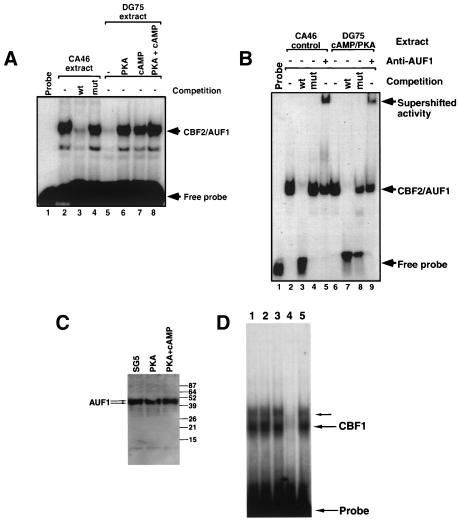
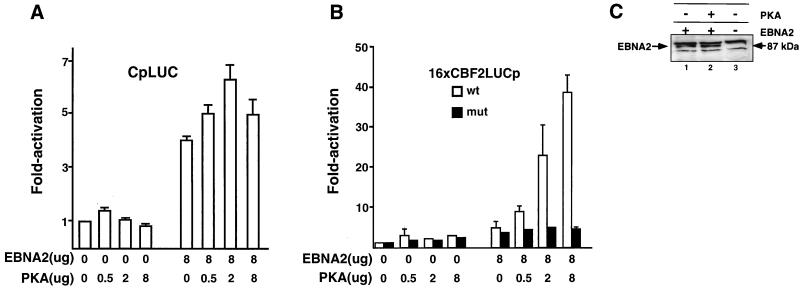
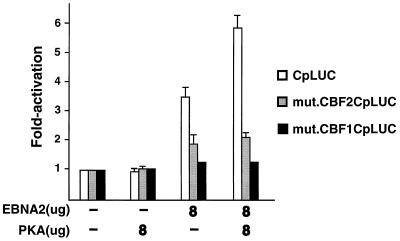
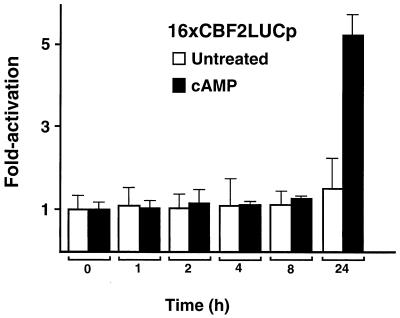
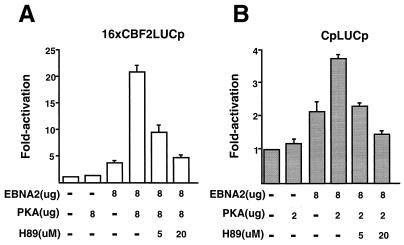
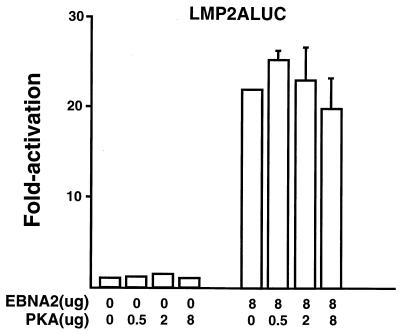
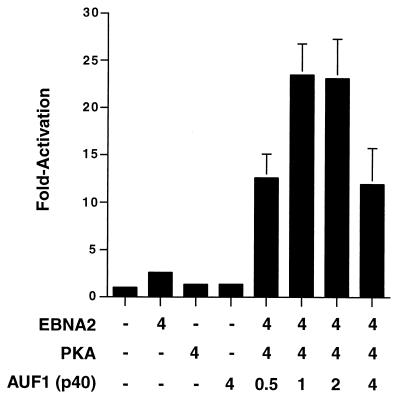
EBNA2 is an Epstein-Barr virus (EBV)-encoded protein that regulates the expression of viral and cellular genes required for EBV-driven B-cell immortalization. Elucidating the mechanisms by which EBNA2 regulates viral and cellular gene expression is necessary to understand EBV-induced B-cell immortalization and viral latency in humans. EBNA2 targets to the latency C promoter (Cp) through an interaction with the cellular DNA binding protein CBF1 (RBPJk). The EBNA2 enhancer in Cp also binds another cellular factor, C promoter binding factor 2 (CBF2), whose protein product(s) has not yet been identified. Within the EBNA2 enhancer in Cp, we have previously identified the DNA sequence required for CBF2 binding and also determined that this element is required for efficient activation of Cp by EBNA2. In this study, the CBF2 activity was biochemically purified and microsequenced. The peptides sequenced were identical to the hnRNP protein AUF1. Antibodies against AUF1 but not antibodies to related hnRNP proteins reacted with CBF2 in gel mobility shift assays. In addition, stimulation of the cellular cyclic AMP (cAMP)/protein kinase A (PKA) signal transduction pathway results in an increase in detectable CBF2/AUF1 binding activity extracted from stimulated cells. Furthermore, the CBF2 binding site was able to confer EBNA2 responsiveness to a heterologous promoter when transfected cells were treated with compounds that activate PKA or by cotransfection of plasmids expressing a constitutively active catalytic subunit of PKA. EBNA2-mediated stimulation of the latency Cp is also increased in similar cotransfection assays. These results further support an important role for CBF2 in mediating EBNA2 transactivation; they identify the hnRNP protein AUF1 as a major component of CBF2 and are also the first evidence of a cis-acting sequence other than a CBF1 binding element that is able to confer responsiveness to EBNA2.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
